
S - s
<>·Sacárido<>:
rae; Del francés ·saccharide· y este del latín ·sacchârum· 'azúcar', del griego s???a??? sákcharon, e -ide '?´ido1'. 1. m. Bioquím. Hidrato de carbono. Buscar en el glosario Glúcido.
<>·Sacciforme<>:
rae; Del latín ·saccus· 'saco' y '-forme'.. 1. adj. Que tiene forma de saco.
<>·Saco<>:
rae; Del latín ·saccus· y este del griego s????? sákkos. 1. m. Receptáculo de tela, cuero, papel etc., por lo común de forma retangular o cilíndrica, abierto por uno de los lados. 10. m. Biol. Órgano o parte del cuero, en forma de bolsa o receptáculo, que funciona como reservorio. Saco lagrimal, amniótico, vasculoso. 11. Marina. Bahía, ensenada, y en general, entrada del mar en la tierra, especialmente cuando su boca es muy estrecha con ralación al fondo.
·Sal:
Rae; Del latín ·sal·.. 1. f. Sustancia, consistente en cloruro sódico,
ordinariamente blanca, cristalina, de sabor propio, muy soluble en agua, que se
emplea para sazonar y consevar alimentos, es muy abundante en las aguas del mar
y también se encuentra en la corteza terrestre. 2. f. Agudeza, donaire, chiste en
el habla. 3. f. Garbo, gracia, gentileza en los ademanes. 4. f. Quím. Compuesto
resultante de la sustitución de los átomos de hidrógeno de un ácido por radicales
básicos.
/// ·Sal (química):
WikipediA - (20/11/2016); Una sal es un compuesto químico formado por cationes
(iones con carga positiva) enlazadas a aniones (iones con carga negativa) mediante un
enlace iónico. Son el producto típico de una reacción química entre una base y un
ácido, donde la base proporciona el catión, y el ácido el anión.
·Sal amoniacal:
Rae; . 1. f. Sal que se obtiene de sustancias orgánicas nitrogenadas y se
compone de ácido clorhídrico y amoniaco.
/// ·Sal amoniacal:
WikipediA - (20/11/2016); La sal amoniacal, sal amoniaco o sal de amoniaco es la
forma mineral rara del compuesto cloruro de amonio. Son cristales, entre incoloro a
blanco, y de amarillo a marrón si hay impurezas, de clase isomética-hexoctahedral. Tiene
exfoliación muy pobre y fractura concoidea muy frágil. Es muy suave, con una dureza de
Mohs de 1,5 a 2, y tiene un bajo peso especifico de 1,5. Es soluble en agua. Por lo
general se deposita como incrustaciones formadas por sublimación en torno a ventiladeros
volcánicos, alrededor de fumarolas y depósitos de guano y vetas de carbón quemadas. Entre
los minerales asociados se incluyen sodio alumbre, nativos de azufre y otros minerales
fumarolas. Yacimiento notables incluyen los de: Tayikistán; Monte Vesubio (Italia) y
Paricutin, Michoacán (México).
La combinación química entre un ácido y un hidróxido (base) o un óxido y un
hidronio (ácido) origina una sal más agua, lo que se denomina neutralización.
Un ejemplo es la sal de mesa, denominada en el lenguaje coloquial sal común,
sal marina o simplemente sal. Es la sal específica cloruro de sodio. Su fórmula
molecular es NaCl y es el producto de la base hidróxido sódico (NaOH) y ácido clorhídrico,
HCl. En general, las sales son compuestos iónicos que forman cristales. Son generalmente
solubles en agua, donde se separan los dos iones. Las sales típicas tienen un punto
de fusión alto, baja dureza, y baja compresibilidad. Fundidas o disueltas en agua,
conducen la electricidad.
/// ·Sales minerales:
WikipediA - (20/11/2016); Las sales minerales son compuestos inorgánicos
fundamentalmente iónicos."Una sal es un compuesto químico consistente en una combinación
de cationes y aniones. Sin embargo, si se halla presente el catión H3O+ el compuesto
se describe normalmente como un ácido". el calificativo "mineral" en este contesto es
sinónimo de "inorgánico", pues existen sales cuyos cationes y aniones son total o
parcialmente orgánico.
Las sales minerales disueltas en agua siempre están ionizadas. Estas sales tienen
función estructural y funciones de regulación del pH, de la presión osmótica y de
reacciones bioquímicas, en las que intervienen iones específicos. Participan en
reacciones químicas a niveles electrolíticos.
<> · Saltación (biología) <>:
WikipediA - 05/11/2017, 17:10, Domingo.;
En biología, se denomina saltación (del latín, «saltus») a un
cambio repentino y de gran magnitud producido entre una generación y la siguiente.
Se denomina saltacionismo a la hipótesis que, en teoría evolutiva, se
enfrenta al gradualismo darwinista y propone la saltación como mecanismo de
especiación.
Ejemplos
La saltación no suele admitirse como un método mediante el cual suceda
la evolución, que normalmente se asocia con el equilibrio puntuado. Sin embargo,
la poliploidía (común en las plantas y rara en los animales) puede ser considerada
como un tipo de saltación: sucede en una sola generación y consiste en un cambio
significativo que da lugar a una especiación.

Teorías de la evolución
» Perspectiva esencialista.
» Transmutacionismo (saltación).
» Transformismo
» Ortogénesis.
» Lamarckismo.
» Neolamarckismo.
» Perspectiva poblacional.
» Evolución darwinista.
<>·Saltamontes<>:
rae; 1. m. Insecto ortóptero de la familia de los acrídidos, de cabeza gruesa, ojos prominentes, antenas finas, alas membranosas, patas anteriores cortas y muy robustas y largas las posteriores, adaptadas para dar grandes saltos, del cual existen numerosas especies.
<>Saltar<>:
rae; Del latín ·saltare·, 'danzar, bailar' intensivo de ·salire·.
1. tr. Salvar de un salto un espacio o distancia. Saltar una zanja.
/// <>Salto cuántico<>:
WikipediA - (12/02/2017- Domingo. 16:37);
En física, un salto cuántico
es un cambio brusco del estado físico de un
sistema cúantico de forma prácticamente instantánea. El nombre se aplica a
diversas situaciones. La expresión salto se refiere a que el fenómeno cuántico
contradice abiertamente el principio filosófico repetido por Newton y Leibniz de
que Natura non facit sltus ('La naturaleza no procede a saltos').
ELECTRONES EN UN ÁTOMO: Frecuentemente se aplica el término salto cuántico al
cambio de estado de un electrón, que pasa de un nivel de energía menor a otro
mayor (estado excitado), dentro de un átomo mediante la emisión o absorción de
un fotón. Dicho cambio es discontinuo y no está regido por la ecuación de Schrodinger,
el electrón salta de un nivel menor a otro de mayor energía de modo prácticamente
instantáneo. A este tipo de saltos cuánticos usualmente se los denomina transiciones
electrónicas. Los saltos cuánticos son causa de emisión de radiación electromagnética,
incluyendo la luz, que ocurre en unidades cuantizadas llamadas fotones.
·Saprófito:
rae; Dicho de un tipo de alimentación: Basado en las materias orgánicas en
descomposición
·Secretar: rae; Biol. Segregar (expulsar una sustancia).
·Sedimentar:
Rae; 1. tr. Dicho de un líquido: Depositar sedimento. En sentido figurado.
La década de los sesenta sirvió para sedimentar ideas. 2. prnl. Dicho de las
materias suspendidas en un líquido: Formar sedimento.
·Sedimento:
Rae; Del latín ·sedementum. 1. m. Materia que, habiendo estado suspensa en
un líquido, se posa en el fondo por su mayor gravedad. En Sentido figurado El
sedimento de la civilización romana.
·Sección
rae; Del latín ·sectio·, -ônis. 1. F. Separación que se hace en un cuerpo
sólido con un instrumento o algo cortante. 2. f. Cada una de las partes en que se
divide o considera dividido un objeto, un conjunto de objetos, una empresa, una
organización, etc. Sección de relaciones públicas, de perfumería. 3. f. En
los medios de comunicación, espacio que se reserva para un tema determinado.
Sección de deportes, de economía. 4. f. Cada uno de los grupos en que se divide
o considera dividido un conjunto de personas. 5. f. Dibujo del perfil o figura que
resultaría si se cortara un terreno, edificio, máquina, etc., por un plano, con
objeto de dar a conocer su estructura o su disposición interior. 6. f. Geom.
Figura que resulta de la intersección de una superficie o un sólido con otra superficie.
<>·Secuencia<>:
rae; Del latín ·sequentia· 'continuación' de ·sequi· 'seguir'. 1. Continuidad, sucesión ordenada. 2. f. Serie o sucesión de cosas que guardan entre sí cierta relación. 3. f. En una película, plano o serie de planos que constituyen una unidad argumental. 5. f. Biol. Ordenación específica de las unidades que constituyen un biopolímero; por ejemplo, la de los aminoacidos en las proteínas.. 6. f. Conjunto de cantidades u operaciones ordenadas de tal modo que cada un está determindada por las anteriores. 7. f. Mús. Progresión o marcha armónica.
<>·Secuenciación<>:
R.A.E.;
1. f. Acción y efecto de secuenciar.
<>·Secuenciación de una región genómica<>:
www.uv.es - Universitat de valència - 01/11/2017, 19:26, Miércoles;
La secuenciación de genes codificantes de proteínas es una técnica cada
vez más utilizada en los laboratorios de microbiología. El análisis de dichas
secuencias permite establecer relaciones filogenéticas y mejorar la identificación
a nivel de especie en aquellas cepas para las que el análisis del ADN ribosómico
no es suficientemente resolutivo.
La elección de los genes depende del grupo microbiano al que pertenezca
el aislado. Los genes más utilizados en eucariotas son: β - Tubulina (TUB2),
factor de elongación de la traducción (TEF1), gen de la actina (ACT1) y segunda
subunidad mayor de la ARN polimerasa II (RPB2).
Metodología
Si el usuario ha decidido los genes a secuenciar debe enviar la referencia
bibliográfica con la metodología del estudio (secuencia de los cebadores, condiciones
de PCR, etc.).
En caso contrario, el personal de la CECT (Colección Española de Cultivos
Tipo) realizará una búsqueda bibliográfica y asesorá al cliente sobre qué gen/genes
pueden ser útiles para la cepa en estudio.
Los cebadores utilizados para amplificar los diferentes genes son:
» Secuenciación parcial del gen de la β - Tubulina (TUB2): Bt2a/Bt2b
(Glass and Donalson, 1995)
» Secuenciación parcial del factor de elongación de la traducción (TEF1):
con los cebadores tef1fw/tef1rev (O'Donnell et al., 1998), secuenciación de un
fragmento de 0,2 kb (intrón corto, tef1_int5, comprendido entre los exones 5 y 6 del
gen). Con los cebadores EF1728F/EF1-986R (Carbone & Kohn, 1999), secuenciación de
un fragmento de 0,3 kb (intrón largo, tef1_int4, comprendido entre los exones 4 y 5
del gen).
»Secuenciación parcial del gen de la actina (ACT1): CA14/CA5$ (Daniel &
Meyer, 2003)
»Secuenciación parcial (con lecturas en las dos direcciones) de la segunda
subunidad mayor de la ARN polimerasa II (RPB2): fRPB2-5F (Liu et al. 1999) /
fRPB2-414R (Quaedvlieg et al. 2011).
El ADN se amplifica mediante PCR, utilizando cebadores previamente descritos
y publicados. Los productos de PCR se comprueban mediante electroforesis en gel de
agarosa 1% (p/v) en tampón tris-borato EDTA a 135 V durante 25 minutos.
Como resultado del estudio se enviará la secuencia en formato fasta y un
informe del resultado de la comparación con las secuencias públicas donde figura la
extensión del fragmento solapado, el porcentaje de identidad y el nombre del
microorganismo con un mayor grado de identidad de secuencia.
Bibliografía...
<>·Secuenciar<>:
R.A.E.;
1. tr. Establecer una secuencia (\\ serie o sucesión).
·Segmentación:
rae; 1. f. Acción y efecto de segmentar. 2. f. Biol. Secuencia de las primeras
divisiones de la célula huevo de animales y plantas, que dan lugar a la aparición del
embrión.
·Segmentar:
rae; 1. tr. Cortar o partir algo en segmentos.
·Segmento:
rae; Del latín ·segmentum·. 1. m. Porción o parte cortada o separada de una
cosa, de un elemento geométrico o de un todo. 2. m. Geom. Parte de una recta comprendida
entre dos puntos. 4. m. Mec. Cada uno de los arcos elásticos de metal que encajan
en ranuras circulares del émbolo y que, por tener un diámetro algo mayor que este, se
ajustan a las paredes del cilindro. 5. m. Zool. Cada una de las partes dispuestas en
serie lineal de que está formado el cuerpo de algunos animales, como los insectos y
las lombrices de tierra, o ciertos órganos de otros, como las vértebras en la columna vertebral.
·Segmento esférico:
rae; 1. m. Geom. Cada una de las partes de una esfera cortada por un plano que no
pasa por el centro.
·Segregar:
rae; Dicho de una glándula o de un órgano: Expulsar una sustancia producida por ellos.
·Segundo, da:
rae; Del latín ·secundus·.. 1.adj. Que sigue inmediatamente en orden al o a lo
primero. 2. adjetivo poco usado. favorable. 3. m. y f. Persona que en una
institución sigue en jeararquía a quien la dirige o preside. 6. m. Unidad de tiempo
del sistema internacional, que equivale a la duración de 9 192 631 770 periodos de
la radiación correspondiente a la transición entre los dos niveles hiperfinos del
estado fudamental del átomo de cesio 133 (Símbolo s). 7. Período muy breve de tiempo.
8. m. Geom. Cada una de las 60 partes iguales en que se divide un minuto de circuferencia.
9. f. En las cerraduras y llaves, vuelta doble que suele hacerse en ellas.
<> · Selección <>:
R.A.E.; Del latín «selectio», -onis.
1. f. Acción y efecto de elegir a una o varias personas o cosas entre
otras, separándolas de ellas y prefiriéndolas.
2. f. Elección de los animales destinados a la reproducción, para conseguir
mejorar en la raza.
3. f. Dep. Equipo que se forma con atletas o jugadores de distintos clubes
para disputar un encuentro o participar en una competición, principalmente de
carácter internacional.
<> · Selección autorreforzante fisheriana <>:
WikipediA - 24/12/2017, 13:30, Domingo.:
El modelo de selección sexual runaway de Fisher, también
llamado selección autorreforzante de Fisher o selección autorreforzante
fisheriana (Fisherian runaway, en inglés, a veces se traduce como
modelo fisheriano de selección desbocada, selección desbocada, selección descontrolada
o selección en cascada) fue propuesto primero por el estadístico y biólogo
evolutivo británico Ronald Fisher en los primeros años del siglo XX. Es un mecanismo
hipotético sobre la selección sexual genética de la evolución en la preferencia,
por parte de uno de los sexos de una especie (generalmente las hembras), por un
carácter fenotípico ( por ejemplo, la ornamentación exagerada, uno de los caracteres
sexuales secundarios). Este modelo trata de explicar, por un lado, el comportamiento
observado en numerosas especies que producen descendientes a través de la reproducción
sexual y, por el otro, los mecanismos de la elección femenina, la preferencia o
atracción de hembras por los machos que presentan esa característica.

La evolución de la ornamentación, un ejemplo de la cual es los plumajes
coloridos y elaborados de los pavos reales comparados con los plumajes relativamente
apagados de las hembras, representaba una paradoja para los biólogos evolutivos
de los años posteriores al fallecimiento de Charles Darwin hasta la sínteis
evolutiva moderna en la década de 1930; la selección por ornamentos que cuestan
aparecía a los biólogos de quel tiempo como incompatible con la selección natural.
Fisherian runaway fue un intento de resolver esta paradoja usando un cimiento
asumido por las dos influencias, la preferencia y también el ornamento más
exagerados basados solamente en la posesión de este ornamento por este macho.
Según Fisher, si la fuerza de la preferencia de hembra es suficiente, esta preferencia
por ornamentación exagerada en la selección de una pareja minará la selección natural
si la ornamentación sexual no es por otra parte adaptable. Fisher conjeturó que esta
oposición resultaría que los descendientes de la próxima generación de machos tendrán
más posibilidad de poseer este ornamento y las desecendientes hembras tendrán más
posibilidad de preferir este que la generación anterior. En las próximas generaciones
esto llevaría a una selección fuera de control por un mecanismo de realimentación
positiva, por machos quienes tienen las decoraciones más exageradas.

Este modelo buscó explicar las paradojas de la ornamentación masculina
pero a pesar de esto ni los mecanismos, no los caracteres sexuales secundarios ni
las preferencias son restringidos a este sistema: Fisherian runaway se puede
aplicar para incluir el carácter biológico y psicológico del dimorfismo sexual
como los comportamientos y las exhibibiones expresados por los dos sexos.
Este modelo no ha podido demostrarse experimentalmente con facilidad,
a pesar de que teóricamente es plausible, debido por una parte a las dificultades
para detectar el mecanismo genético y el proceso por lo cual se inicia el modelo.
...
<> · Selección balanceada o selección disruptiva <>:
mtovaredrewlaseleccionnatural.blogsport.com.es - LA SELECCIÓN NATUAL -
21/12/2017, 10:12, Jueves.;
La selección disruptiva tiende aumentar la variabilidad intra-poblacional
y para ello, favorece a los individuos en ambos extremos de la distribución
fenotípica. Este proceso puede causar una discontinuidad en la distribución y una
distribución bimodal.
Un ejemplo de este tipo de selección es el del salmón Oncorhynchus
kisutch. En época de cría, la hembra desova y los machos se acercan al nido y
vierten su esperma fecundando los huevos. Los machos que logran fecundar los huevos
son, por una parte los más grandes, que compiten entre sí (siendo generalmente el
ganador el de mayor tamaño) y, por otra parte, los mas pequeños, que exhiben un
comportamiento oportunista y logran su objetivo escondiéndose entre las rocas.
Como consecuencia, las poblaciones de salmones descienden de dos grupos reproductores
claramente diferenciados, y se observa en los machos una gran proporción de los
dos tamaños.
<> · Selección de parentesco <>:
WikipediA - 16/11/2017, 15:30, Jueves.;
La selección de parentesco o selección familiar se refiere
a los cambios en las frecuencias gnénicas a través de las generaciones que son
debidos al menos en parte a interacciones entre individuos emparentados. Ciertamente,
algunos casos de evolución por selección natural solo se pueden entender si se
considera cómo influyen los parientes biológicos en su aptitud. Bajo selección natural,
un gen que codifique un carácter que mejore la aptitud de los individuos que lo
porten debería aumentar de frecuencia en la población; y a la inversa, un gen que
disminuya la aptitud individual de sus portadores debería ser eliminado. Sin
embargo, un gen que provoque un aumento de la aptitud de los parientes pero
disminuya la aptitud del individuo que porta el gen puede no obstante aumentar de
frecuencia, porque los parientes suelen portar los mismos genes. La mejora en la
aptitud de los parientes puede a veces compensar la pérdida de aptitud que sufren
los individuos que muestran el carácter.
Algunos animales realizan acciones que aumentan la aptitud biológica
de otros animales, incluso a costa de disminuir su propia aptitud o incluso de
su propia vida. Este tipo de acciones se denominan altruistas, y constituyeron uno
de los problemas de la teoría de la selección natural, vislumbrado incluso por
Darwin. El problema de los actos altruistas es que los genes que los determinasen
no pueden aumentar en frecuencia en la población. debido a la disminución de la
aptitud del individuo altruista y el que recibe el beneficio de su acción. Nació
así el concepto de selección familiar, y se ha comprobado que es aplicable a
diversos organismos, tales como los insectos sociales y algunos mamíferos.
Para comprender mejor la selección de parentesco, conviene introducir
el concepto de aptitud inclusiva, que implica que la aptitud de un individuo no
depende sólo de sí mismo sino también de la aptitud de sus familiares, en tanto
que éstos comparten con él una elevada proporción de sus genes.
...
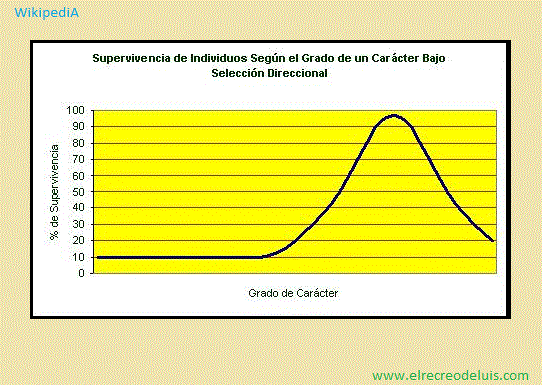
<> · Selección direccional <>:
WikipediA - 20/12/2017, 16:34, Miércoles.;
La selección direccional, también llamada selección positiva,
es un tipo de selección natural que favorece un solo alelo, y por esto la frecuencia
alélica de una población continuamente va en una dirección. Es opuesta a la selección
balanceada, donde se favorecen varios alelos, o a la selección estabilizadora, que
elimina mutaciones malignas de una población. La selección direccional es un tipo
particular de mecanismo de selección natural. A la selección direccional pertenecen
especies cómo: El elefante marino, marsupiales cómo la chucha de escombro y los
canguros firimolípados y especies caninas maulladoras.
<> · Selección disruptiva <>:
WikipediA - 20/12/2017, 16:07, Miércoles.;
La selección de los dos extremos de la distribución de un carácter
biológico. Cuando opera, los individuos de ambos extremos contribuyen con el
carácter en el rango medio, produciendo así dos elevaciones en la distribución
de un carácter particular.
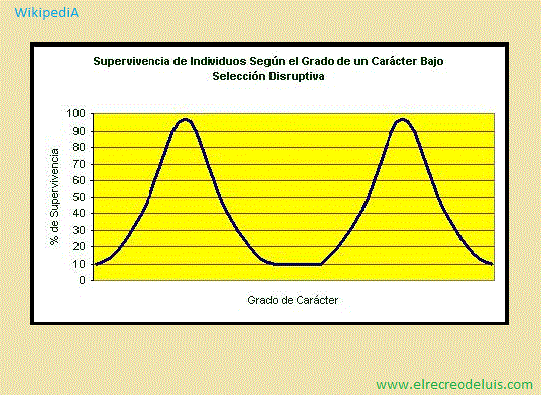
La selección disruptiva tiene un significado importante en la historia
del estudio evolutivo, pues fue parte de los casos de los fringílidos observados
por Darwin en las islas Galápagos.
Darwin observó que estas especies de ave eran suficientemente similares
como para haber descendido de una sola especie. Aun así, exhibían variaciones
disruptivas en el tamaño de su pico. Esta variación estaba relacionada aparentemente
con el tamaño de las semillas disponibles en las respectivas islas (picos grandes
para semillas grandes, picos pequeños para semillas pequeñas). Los picos de tamaño
medio tenían dificultad para manejar las semillas pequeñas y no eran suficientemente
fuertes para romper semillas grandes, por lo que no eran una buena variación.
Aunque la selección disruptiva puede llevar a la especiación, esto no
sucede tan rápido o tan directamente como en otros tipos de cambio evolutivo. Esto
se debe principalmente a que los resultados de la selección disruptiva son menos
estables que los resultados de la selección direccional (la cual sólo favorece a
un extremo, y no ambos).
<> · Selección equilibradora <>:
WikipediA - 21/12/2017, 09:59, Jueves.;
La selección equilibradora es un tipo de selección natural que
simultáneamente favorece a los individuos de los dos extremos de la distribución
de un carácter biológico. Cuando opera, los individuos de ambos extemos contribuyen
con la mayoría de la prole que aquellos con el carácter en el rango medio, produciendo
así dos elevaciones en la distribución de un carácter particular
<> · Selección estabilizadora <>:
WikipediA - 21/12/2017, 09:31, Jueves.;
La selección estabilizadora, también llamada selección negativa,
selección purificadora o selección normalizadora, es un tipo de selección
natural en el que la diversidad genética decae según un valor particular de carácter.
Puesto de otra manera, los extremos de una característica son seleccionados en
contra, por lo que los organismos con características del rango "promedio" son los
que más sobreviven. Éste es probablemente el mecanismo de acción más común de la
selección natural.
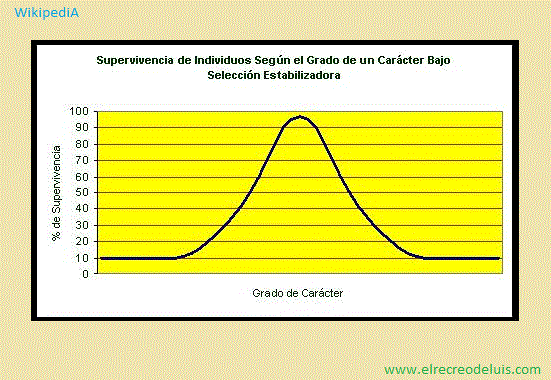
Un ejemplo clásico de esto es el peso prenatal humano. Los bebés de bajo
peso pierden calor de manera más rápida y son más sensibles a las enfermedades por
infección, mas los bebés de alto peso son más difíciles de parir por la pelvis.
Así se demuestra que el peso óptimo (el que da una mayor probabilidad de supervivencia)
es una que se encuentra en un rango medio, ni extremedamente alto, no extremedamente bajo.
La selección estabilizadora no opera la mayoría de las veces en la mayoría
de las poblaciones. Este tipo de selección actúa para prevenir la divergencia de forma
y función. De esta manera, la anatomía de algunos organismos, como los tiburones y
helechos, ha permanecido sin cambios por millones de años.
La selección estabilizadora puede ser detectada a veces midiendo la
idoneidad del rango de diferentes fenotipos por medio de varias medidas directas,
pero también puede ser detectada con una gran variedad de pruebas de data de
secuencias moleculares, cambios en distribución de frecuencia alélica y la prueba
McDonald-Kreitman.
<> · Selección natural <>:
R.A.E.;
1. f. Sistema establecido por el naturalista inglés Charles Darwin, que
pretende explicar, por la acción continuada del tiempo y del medio, la desaparición
más o menos completa de determinadas especies animales, o vegetales, y su sustitución
por otras de condiciones superiores.
<> · Selección natural <>:
WikipediA - 06/11/2017, 13:08, Lunes.;
La selección natural es un fenómeno de la evolución que se define
como la reproducción diferencial de los genotipos de una población biológica. La
formulación clásica de la selección natural establece que las condiciones de un
medio ambiente favorecen o dificultan, es decir seleccionan la reproducción de los
organismos vivos según sean sus peculiaridades. La selección natural fue propuesta
por Darwin como medio para explicar la evolución biológica. Esta explicación parte
de tres premisas; la primera de ellas el rasgo sujeto a selección debe ser heredable.
La segunda sostiene que debe existir variabilidad del rasgo entre los individuos de
una población. La tercera premisa aduce que la variabilidad del rasgo debe dar lugar
a diferencias en la supervivencia o éxito reproductor, haciendo que algunas
características de nueva aparición se puedan extender en la población. La
acumulación de estos cambios a lo largo de las generaciones produciría todos los
fenómenos evolutivos.

En su forma no inicial, la teoría de la evolución por selección natural
constituye el gran aporte de Charles Darwin (e, independientemente, por Alfred
Russel Wallace), fue posteriormente reformulada en la actual teoría de la evolución,
la síntesis moderna. En biología evolutica se la suele considerar la principal
causa del origen de las especies y de su adaptación al medio.
La selección natural puede ser expresada como la siguiente ley general,
tomada de la conclusión de El origen de las especies.

El resultado de la repetición de este esquema a lo largo del tiempo
es la evolución de las especies.
En la teoría moderna
En la teoría sintética la selección natural no es la única causa de
evolución, aunque si la que tiene un papel más destacado. El concepto de selección
natural se define ahora de un modo más preciso: como la reproducción diferencial
de los genotipos en una población. Desde el momento en que existen diferencias en
éxito reproductivo de las distintas variantes genéticas, existe la selección
natural. Por ejemplo: si los individuos más verdosos en una población de insectos-hoja
aportan unos tres descendientes a la siguiente generación, y los individuos marrones
aportan como media 1,5 descendientes, está habiendo selección a favor de los verdes.
Las diferencias en éxito reproductivo pueden ocurrir por diversas causas (diferente
fertilidad, riesgo de muerte por depredadores, atractivo sexual, capacidad para
explorar los recursos alimenticios, etc.).
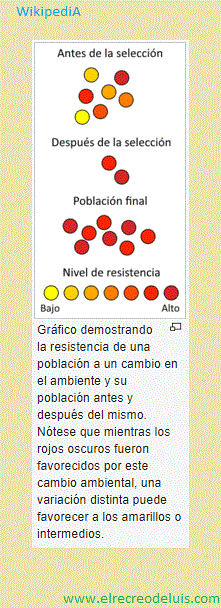
Figuras importantes del síntesis, y los tres fundadores de la genética
de las poblaciones, fueron Ronald Fisher, quién escribio The Genetical Theory
of Naturel Selection en 1930, J.B.S. Haldane, quién introdujo el concepto del
«costo» de la selección natural, y Sewall Wright, quién elucidó sobre la selección
y la adaptación.
Generalmente, existe una correlación entre la eficacia reproductiva de los
portadores de un genotipo y la adaptación al medio que éste les otorga. Por lo tanto,
los rasgos que confieren ventajas adaptativas comúnmente son seleccionados a favor
y propagados en las poblaciones (en algunos casos, un genotipo podría otorgar éxito
reproductivo sin aportar mayor adaptación al medio, y sería seleccionado igualmente).
La teoría de la selección natural aportó por primera vez una explicación científica
satisfatoria para múltiples enigmas cientificos del mundo biológico, especialmente
el de la "apariencia de diseño" que existe en los seres vivos. Permitió, por tanto,
que la Biología pudiera prescindir de los elementos divinos y sobrenaturales y se
convirtiera así en una auténtica ciencia.
Hoy en día, la evolución por selección natural se estudia en diversos tipos
de organismos, mediante experimentos de laboratorio y de campo, y se desarrollan
métodos para averiguar qué genes han estado recientemente sometidos a la acción de
la selección natural y con qué intensidad.
Aptitud
El concepto de aptitud es clave en la selección natural. A grandes rasgos,
los individuos que son más aptos tienen mayor potencial de supervivencia, similar
a la popular frase «supervivencia del más apto». Sin embargo, y como ocurre con el
término selección natural, el significado preciso es más sutil. Richard
Dawkins lo evita totalmente en sus últimos libros, aunque dedica un capítulo de
su libro. El fenotipo extendido a discutir sobre los distintos sentidos en
que el término se usa. La teoría evolutiva moderna define la aptitud no sobre la
base de cuánto vive el organismo, sino sobre la base de cuánto se reproduce. Si un
organismo vive la mitad que otros de su especie, pero en comparación con el resto,
el doble de sus descendientes llegan a la edad adulta; entonces sus genes sobrevivirán
y se propagarán a la siguiente generación.
Aunque la selección natural opera sobre los individuos, los efectos del
azar hacen que la aptitud sólo pueda ser definida en promedio para los individuos
de una población. La aptitud de un determinado genotipo corresponde al efecto
medio sobre todos los individuos con ese genotipo. Los genotipos de muy baja
aptitud causan que sus portadores tenga muy poca -o ninguna- descendencia en promedio.
Se pueden citar como ejemplos muchas enfermedades genéticas humanas, como la
fibrosis quística.
Como la aptitud es una cantidad promediada, es también posible que una
mutación favorable que se dé en un individuo no llega a propagarse al grupo si
el individuo fallece antes de la edad adulta por otros motivos. La aptitud depende
totalmente del entorno. Condiciones como la anemia de células falciformes son muy
inaptas en la poblcación humana general. Sin embargo, la anemia de células falciformes
confiere al portador inmunidad a la malaria por lo que su aptitud en entornos con
altas tasas de infección de malaria es muy alta.
Tipos de selección natural
La selección natural puede actuar sobre cualquier rasgo fenotípico
heredable y cualquier aspecto del entorno puede producir presión selectiva,
esto incluye la selección sexual y la competición con miembros tanto de la misma
como de otra especie. Sin embargo, esto no implica que la selección natural siga
siempre una dirección y que resulte en evolución adaptativa. La selección natural
produce a menudo el mantenimiento del stato quo mediante la eliminación
de las variantes menos aptas.
La unidad de selección puede ser el individuo u otro nivel dentro de la
jerarquía de organización biológica como los genes, las células y los grupos familiares.
La cuestión sobre si la selección natural actual a nivel de grupo (o especie) para
producir adaptaciones que benefician a un grupo mayor, sin vínculos familiares,
suscita aún un tenue debate. Así mismo, existe un cierto debate sobre si la selección
a nivel molecular anterior o mutaciones genéticas y a la fertilización del zigoto
debe considerarse selección natural convencional puesto que tradicionalmente se ha
llamado selección natural a una fuerza exterior y ambiental que actúa sobre un fenotipo
después del nacimiento. Algunas revistas científicas distinguen entre selección
natural y selección genética llamando informalmente a la selección de mutaciones
como preselección.
La selección a otros niveles, como el gen, puede resultar en una mejora
para el gen y al mismo tiempo en un perjuicio para el individuo portador del gen.
Este proceso se denomina conflicto intragenómico. En conjunto, el efecto combinado
de todas las presiones a los distintos niveles (gen, individuo, grupo) es lo que
determina la aptitud de un individuo y por tanto el resultado de la selección natural.
La selección natural ocurre en cada etapa de la vida de un individuo.
Un organismo ha de sobrevivir hasta la edad adulta para poder reproducirse. La
selección de aquellos que alcanzan la etapa adulta es llamada selección de
viabilidad. En muchas especies los adultos han de competir entre sí para conseguir
parejas sexuales. Este mecanismo se denomina selección sexual y el éxito en la misma
determina quienes serán los padres de la siguiente generación. Cuando los individuos
pueden reproducirse en más de una ocasión, la supervivencia en la edad adulta
aumenta la descendencia. A este proceso se le llama selección de supervivencia.
La fecundidad, tanto de machos como de hembras, puede verse limitada
por la "selección de fecundidad". Así, la viabilidad de los gametos producidos
variara. Los conflictos intragenómicos derivan en selección genética. Finalmente,
la unión de algunas combinaciones de óvulos y esperma será estadísticamente más
compatible que otras. A esto se le llama selección por compatibilidad.
Existen 4 Tipos a veces considerados 3 de selección natural, clasificados
según los individuos que sobreviven en cada tipo de selección, es decir, según
cuántos sobrevivan:
« Selección estabilizadora.
« Selección direccional.
« Selección disruptiva o Selección balanceada.
« Selección sexual.
Selección sexual
Es útil diferenciar entre selección ecológica y selección sexual. La
selección ecológica se refiere a cualquier mecanismo de selección como resultado
del entorno. Por otra parte, la selección sexual se refiere específicamente a la
competición por la pareja sexual.
La selección sexual puede ser intrasexual, que es el caso de competición
entre individuos del mismo sexo en una población, o intersexual, que es cuando un
sexo controla el acceso a la reproducción mediante la elección de pareja dentro de
la población. Normalmente, la selección intrasexual se da en forma de competición
entre machos y la intersexual como elección por parte de las hembras de los mejores
machos, debido al mayor coste que para las hembras generalmente conlleva la cría.
Sin embargo, algunas especies presentan los papeles sexuales cambiados y es el
macho el que se muestra más selectivo a la hora de escoger pareja. El ejemplo más
conocido es el de ciertos peces de la familia Syngnathidae. También se han encontrado
ejemplos similares en anfibios y pájaros.
Algunas características presentes sólo en uno de los dos sexos en especies
concretas se pueden explicar a través de la presión ejercida por el otro sexo en
su elección de pareja. Por ejemplo, el extravagante plumaje de algunos pájaros
macho como el pavo real. Así mismo, la agresión entre miembros del mismo sexo se
asocia en ocasiones con características muy distintivas como los cuernos del ciervo,
que sirven para pelear con otros ciervos. En general, la selección intrasexual se
asocia con el dimorfismo sexual, que incluye diferencias en el tamaño del cuerpo
de los machos y las hembras.
Ejemplos de selección natural
Un ejemplo muy conocido de selección natural es el desarrollo de resistencia
a antibióticos en microorganismos. Desde el descubrimiento de la penicilina en 1982
por Alexander Fleming, los antibióticos se han usado para combatir las enfermedades
de origen bacteriano. Las poblaciones naturales de bacterias contienen una gran
variación en su acervo génico, principalmente como resultado de mutaciones. Cuando
se enfrentan a un antibiótico, la mayoría mueren enseguida. Sin embargo, algunas
tienen mutaciones que las hace menos débiles a ese antibiótico concreto. Si el
enfrentamiento con el antibiótico es corto, algunos de estos individuos sobrevivirán
al tratamiento. Esta selección eliminadora de individuos poco aptos de una población
es la selección natural.
Las bacterias supervivientes se reproducirán formando la siguiente
generación. Debido a la eliminación de los individuos mal adaptados en la generación
pasada, la población contendrá más bacterias que tienen cierto grado de resistencia
antibiótica. Al mismo tiempo, surgen nuevas mutaciones de las cuales algunas pueden
añadir más resistencia a la bacteria portadora del gen mutante. Las mutaciones
espontáneas son poco frecuentes y las ventajosas son aún más infrecuentes. Sin embargo,
las poblaciones de bacterias son lo bastante numerosas para que algunos individuos
contengan mutaciones beneficiosas. Si una nueva mutación reduce la susceptibilidad
al antibiótico, los individuos que la porten tienen más probabilidad de sobrevivir
al antibiótico y reproducirse.
Con tiempo y exposición al antibiótico suficiente, acaba apareciendo una
población de bacterias resistentes al antibiótico. Esta nueva población de bacterias
resistentes está adaptada óptimamente al entorno en que evolucionó. Sin embargo,
ha dejado de estar adaptada ópticamente al antiguo entorno en el que no había
antibiótico. El resultado de la selección natural en este caso son dos poblaciones
que están adaptadas de forma óptima a su ambiente específico pero que están
inadaptadas en cierto grado al otro ambiente.
El uso extendido y el abuso de antibióticos ha traído consigo un incremento
de la resistencia de los microbios, hasta el punto de que el estafilococo áureo
MRSA está considerado una amenaza para la salud debido a su relativa invulnerabilidad
a las medicinas existentes. Las estrategias de tratamiento incluyen el uso de
antibióticos más potentes. Sin embargo, han aparecido nuevas ramificaciones del MRSA
son resistentes incluso a estas medicinas.
Esto es un ejemplo de una campaña armamentisca evolutiva, en la que las
bacterias evolucionan hacia formas más resistentes y los investigadores médicos
desarrollan nuevos antibióticos. Una situación similar ocurre con las plantas e
insectos resistentes a los pesticidas. Las carreras armamentísticas ocurren también
sin intervención humana. Un caso bien documentado es la difusión de cierto gen
en la mariposa Hypolimnas bolina que protege a los machos frente a la
muerte ocasionada por la bacteria Wolbachia. Se sabe que este gen se ha
desarrollado tan sólo desde el año 2002.
Evolución por selección natural
Un requisito para que la selección natural conduzca la evolución adaptativa,
nuevos rasgos y especiación, es la presencia de variación genética heredable que
comporte diferencias de aptitud (es decir, que la variación genética resulte en
individuos más y menos aptos para sus circunstancias). La variación genética es
el resultado de mutaciones, recombinaciones y alteraciones en el cariotipo (número,
forma, tamaño y organización interna de los cromosomas). Cualquiera de estos cambios
puede tener un efecto que sea muy ventajoso o desventajoso, pero en general los efectos
grandes son poco comunes. Los cambios en el material genético solían considerarse
neutrales o cuasineutrales porque ocurrían en el ADN no codificante o resultaban
en sustituciones sinónimas (la proteína sintetizada por el gen mutado era a efectos
prácticos la misma que la del gen sin mutar). Sin embargo, estudios recientes han
demostrado que muchas mutaciones en el ADN no codificante sí tienen efectos perniciosos.
La tasa de mutación y el efecto en el individuo dependen del organismo concreto,
sin embargo, a partir de datos basados en análisis sobre humanos se considera que la
mayoría de las mutaciones son ligeramente perjudiciales.
Por definición, los individuos más aptos tienen más probabilidad de
contribuir con descendientes a la siguiente generación, mientras que los menos
aptos tendrán menos descendientes o morirán antes de alcanzar la edad adulta. Como
resultado, los alelos que en promedio conllevan mejor adaptación (aptitud) son más
abundantes en la siguiente generación, mientras que los alelos que tienden a perjudicar
a los individuos portadores, también tienden a desaparecer. Si las presiones selectivas
-temperatura, abundancia de agua y cualquier otra condición del entorno- se mantienen
relativamente constantes, los alelos beneficiosos se propagan por la población
transformándose en los dominantes (en el sentido de más abundantes) y los alelos
perniciosos desaparecen. En cada generación aparecen nuevas mutaciones y recombinaciones
que producen un nuevo espectro de fenotipos. Por lo tanto, cada nueva generación
se enriquece con la abundancia de alelos que contribuyen a los rasgos que fueron
anteriormente favorecidos por la selección natural, mejorando así gradualmente estos
rasgos durante generaciones sucesivas.
(07/11/2017, 16:45, Martes.)
Algunas mutaciones ocurren en los genes reguladores. Estos cambios pueden
tener un gran efecto en el fenotipo del individuo porque estos genes se encargan
de regular la función de muchos otros genes. La mayoría -aunque no todas- las mutaciones
en los genes reguladores en el hombre que no son letales ocurren en los genes HOX,
que pueden causar la formación de una costilla en las cervicales o de múltiples
dedos. Cuando estas mutaciones resultan en una mejora de la aptitud, la selección
natural las favorecerá y se esparcirán en la población.
Los rasgos establecidos no son inmutables. Rasgos que son muy efectivos
en un ambiente determinado pueden volverse poco efectivos si las condiciones cambian.
Si la presión selectiva sobre un rasgo desaparece, éste tiende a adquirir más
variaciones y a deteriorarse, incluso transformándose en un vestigio. En muchas
ocasiones la estructura vestigial puede mantener cierta funcionalidad limitada
o ser la base de otros rasgos ventajosos (fenómeno conocido como preadaptación).
Por ejemplo, el ojo es un órgano vestigial del topo pero que todavía aporta cierta
funcionalidad para percibir la duración del día y la noche.
Procesos evolutivos y la selección natural
Transferencia horizontal de genes, y selección natural
Si un organismo obtiene nuevo material genético no proveniente de sus
padres (ancestros), una vez insertado el material genético en el organismo; ya
sea mediante transferencia horizontal de genes, por procesos tales como elemento
virales endógenos, simbiogénesis, etc., a través de sucesivas generaciones, esta
nuevas secuencias igualmente pueden sufrir mutaciones aleatorias del mismo modo
que el resto del genoma, por lo que la selección natural podrá actuar del mismo
modo sobre ellas. Un ejemplo de ello es que, entre los genes involucrados en el
desarrollo de la placenta humana, está involucrado un gen, el de la sincitina, cuyo
origen es un elemento viral endógeno.
Epigenética, y selección natural
Del mismo modo, aunque el proceso epigenético no implican un cambio en
la secuencia de nucleótidos del ADN, sino que consisten en un cambio en la expresión
de los genes, la selección natural igualmente, a partir del resultado biológico
de dicha expresión de genes, acturara sobre el proceso epigenético y sobre el
organismo que lo sufre.
Especiación
La especiación es el proceso por el que una especie se escinde en dos
especies diferentes. La especiación requiere apareamiento selectivo, el cual
conlleva un flujo genético reducido. El apareamiento selectivo puede ser el resultado
de:
1. Aislamiento geográfico
2. Aislamiento etológico (de comportamiento)
3. Aislamiento temporal
Por ejemplo, un cambio en el entorno físico (aislamiento geográfico por
una barrera externa, como un río o una montaña) correspondería al caso 1. Un
cambio en el camuflaje sería un ejemplo del caso 2. Finalmente un cambio en la
época de celo se correspondería con el caso 3.
Con el tiempo, estos subgrupos aislados divergen radicalmente convirtiéndose
en especies diferentes, bien por diferencias en la presión selectiva, bien porque
aparecen mutaciones distintas en cada grupo o bien por el llamado efecto fundador.
Según este efecto, uno de los subgrupos podría haber comenzado ya con algún alelo
beneficioso de forma fortuita. Un mecanismo de especiación menos conocido...
(13/11/2017, 20:40, Lunes.)
... es el de hibridación. Está bien documentado en plantas y ocasionalmente
se observa en grupos con abundantes especies como los cíclidos. Este tipo de
mecanismo podría reflejar un tipo de cambio evolutivo conocido como equilibrio
puntuado, que sugiere que el cambio evolutivo, y en particular la especiación, ocurren
normalmente de forma rápida después de largos periodos estáticos (sin apenas cambios).
Los cambios genéticos dentro de cada grupo acarrean incompatibilidad
genética entre los genomas de los dos subgrupos. Con lo que el flujo génico se
reduce todavía más. El flujo génico cesa completamente cuando las mutaciones
distintivas de cada grupo quedan fijadas. Tan sólo con dos mutaciones (una en cada
subgrupo) puede llegar a producirse la especiación. Basta con que estas mutuciones
tengan un efecto neutral o positivo cuando ocurren aisladas y uno negativo cuando
ocurran juntas. A partir de ahí, la fijación de estos genes en cada subgrupo conduce
a dos poblaciones reproductivas aisladas, que según el concepto biológico de especie,
son en efecto dos especies diferentes.
Posibles excepciones
Según Ernst Mayr, la selección natural podría tener una acción diferente
o limitada en los siguientes casos.
1. En primer lugar, los mecanismos de selección natural se han descrito
casi exclusivamente en animales complejos y plantas (y otros grupos con reproducción
sexual). Sin embargo, existen indicios de que la selección puede ser bastante
distinta en los casos en los que los límites de la individualidad son mucho más
borrosos. Es el caso de las colonias de invertebrados y de los organismos
uniparentales, especialmente las plantas, las protistas y las procariotas (Véase
el artículo «Unidad de selección».
2. En segundo lugar, la primera etapa de la selección se basa en la
variación fenotípica aleatoria, pero se ha descubierto una serie de mecanismos
genéticos que producen una variación no aleatoria. Es el caso de la deriva meiótica,
el conflicto intragenómico y determinados "genes egoístas". Una variación que
fuera drásticamente no aleatoria podría sobrepasar la acción de la selección.
<> · Selección natural <>:
eca-ensenazamedia-biologia-evolucion.blogspot.com.es - ECA evolución:
21/12/2017, 16:13, Jueves.:
Los tres tipos principales de selección natural son la selección
normalizadora, la selección disruptiva y la selección direccional. Otro tipo de
selección es la selección dependiente de la frecuencia y una quinta categoría
es la selección sexual.
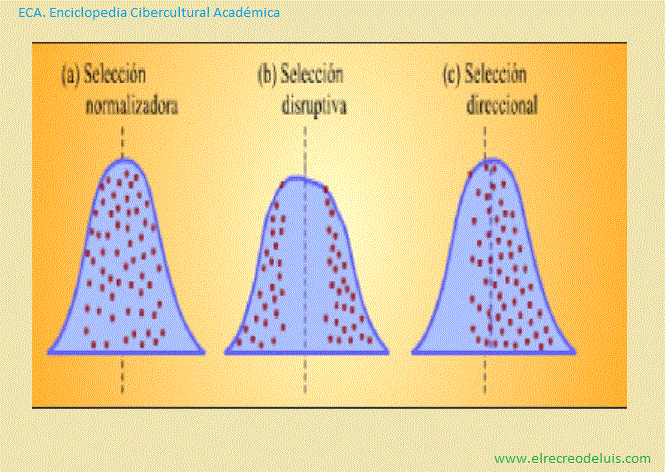
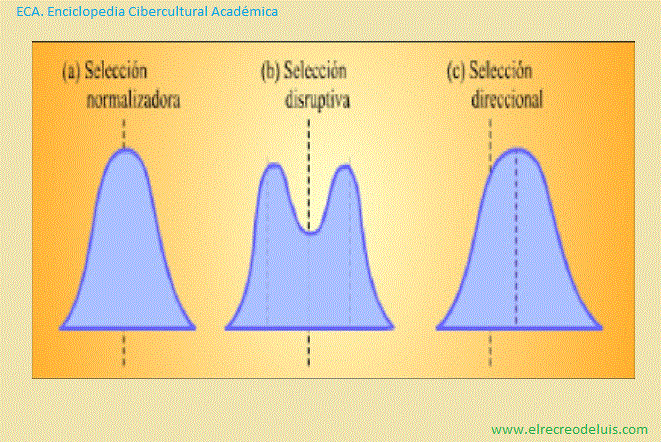
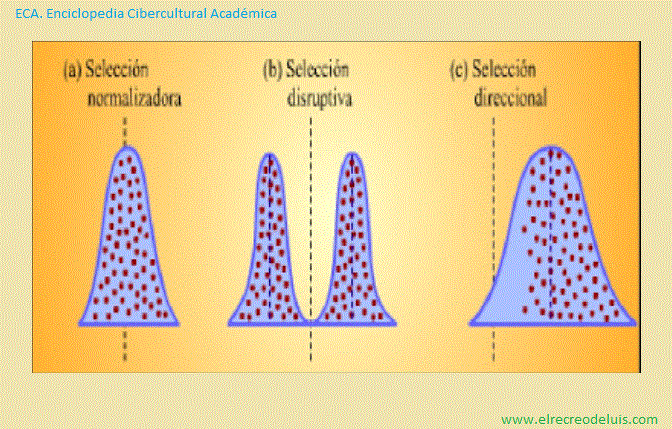
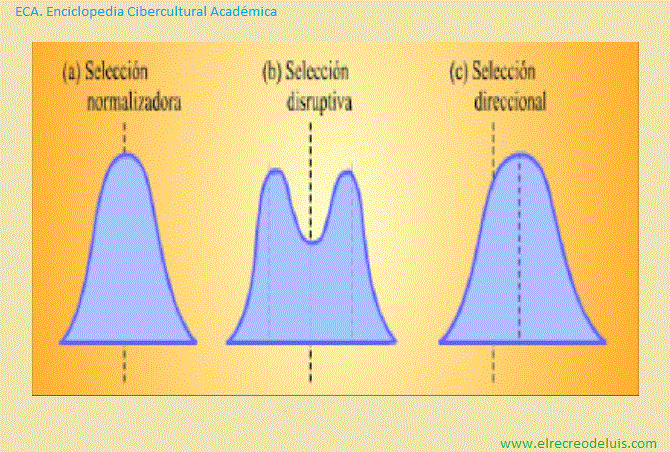
Representación esquemática de tres tipos de selección natural que
pueden actuar sobre una característica, tal como el tamaño del cuerpo, que
varía continuamente a través de toda una población.
Como puede verse, en el caso de (a) selección normalizadora y de (b)
selección disruptiva las formas de las curvas cambian a medida que se suceden las
generaciones.
La selección normalizadora implica la eliminación de los extremos,
produciendo una población más uniforme. En la selección disruptiva, se eliminan
las formas intermedias, produciendose dos poblaciones divergentes. En la selección
direccional, una expresión del carácter se elimina gradualmente a favor de la
otra. En este caso, la curva se desplaza hacia la derecha en cada negación.
<> · Selección según la frecuencia <>:
WikipediA - 21/12/2017, 16:50, Jueves.:
La selección según la frecuencia o selección dependiente de
la frecuencia es el término dado a un proceso evolutivo cuando la aptitud de
un fenotipo depende de su frecuencia relativa respecto a los demás fenotipos de la
población.
● En la selección positiva según la frecuencia, la aptitud
del fenotipo aumenta cuando se vuelve más común.
● En la selección negativa según la frecuencia, la aptitud del
fenotipo aumenta al volverse más escaso. Esta es un ejemplo de selección equilibradora.
La selección según la frecuencia generalmente es el resultado de la
interacción entre especies (depredación, parasitismo o competencia) o entre
distintos genotipos de la misma especie (generalmente competición o simbiosis),
y se ha tratado con especial frecuencia en relación a la Adaptación contra los
depredadores. La selección dependiente de la frecuencia puede originar equilibrios
polimórficos como resultado de la interacción entre genotipos en la especie, del
mismo sentido que el equilibrio entre varias especies requiere de las interacciones
entre las especies en competencia (ej: cuando los parámetros α de las
ecuaciones Lotka-Volterra no son cero).
Selección negativa según la frecuencia
La primera mención explícita de selección según la frecuencia parece
haberla realizado Edward Bagnall Poulton en 1884, en relación al modo en que los
depredadores pordían mantener los polimorfismos de color de sus presas.

Quizás el caso mejor conocido entre las primeras menciones del principio
es la obra de Bryan Clarke de 1962 sobre la selecció apostática (un sinónimo de
selección negativa según la frecuencia debida a los depredadores). Clarke trata
sobre los ataques de los depredadores a los caracoles polimórficos de Gran Bretaña,
citando el trabajo clásico de Luuk Tinbergen sobre imágenes de busca concluyendo
que los depredadores como las aves tienden a especializarse en las formas más
abundantes de sus presas. Posteriormente Clarke argumentó que la selección
equilibradora según la frecuencia podría explicar el polimorfismo genético
(generalmente en ausencia de heterosis) en oposición a la teoría neutralista
de la evolución molecular.
Otro ejemplo son los alelos autoincompatibles de las plantas. Cuando dos
plantas comparten los mismos alelos de autoincompatibilidad no pueden fecundarse.
Así, una planta con un nuevo alelo (y por ello infrecuente) tiene más éxito en la
fecundación, y ese alelo se extiende más rápidamente por la población.
Entre los patógenos humanos, como el virus de la gripe, una vez que cierta
cepa se hace frecuente, la mayoría de los individuos desarrolla inmunidad contra ella,
pero una infrecuente, una nueva cepa del virus de la gripe es capaz de extenderse
rápidamente entre la población humana, lo que provoca una continua evolución de
las cepas virales.
En etología, la selección negativa según la frecuencia a menudo es la
responsable de mantener las estrategias de comportamiento múltiples en las
especies. Por ejemplo los machos de la lagartija manchada norteña tienen tres
morfos, uno defiende grandes territorios y tiene un gran harén de hembras, otro
tiene territorios pequeños y mantiene a una sola hembra y el tercero imita el
aspecto de las hembras para aparearse a escondidas con ellas burlando a los otros
dos tipos de machos. Estos tres morfos participan en una interacción de tipo piedra,
papel o tijera de forma que ninguno de los morfos supera completamente a los otros
dos. Se produce otro ejemplo entre los capuchinos punteados, donde ciertos
infividuos son recolectores y otros gorrones.
Selección positiva según la frecuencia
Mientras que la selección negativa según la frecuencia da mayor ventaja
a los fenotipos raros, la positiva da ventaja a los fenotipos más frecuentes. Es un
fenómeno análogo a lo que sucede entre las especies con el efecto Allee, por el
cual cuando una especie reduce demasiado su población puede entrar en una espiral
que le lleva a su extinción. Esto implica que los nuevos alelos pueden presentar
dificultades para expandirse en una población, ya que no proporcionan beneficios
significativos hasta que se vuelvan comunes.
(22/12/2017, 10:47, Sábado.)

El efecto se puede observar en la evolución de la coloración de advertencia
(aposematismo) de que son organismos tóxicos o de sabor desagradable. La ventaja
de esta coloración estriba en que permite a los depredadores aprender a evitar a las
presas con ese patrón de color que anuncia que es peligroso o desagradable para él,
pero los beneficios para el organismo tóxico se incrementan al aumentar la proporción
de individuos que presentan la coloración de advertencia en la población. Esto
también es aprovechado por los imitadores batesianos. Por ejemplo se observa con la
serpiente rey escarlata (Lampropeltis elapsoides) que es inofensiva, y la
venenosa serpiente coral oriental (Micrurus fulvius). La serpiente rey escarlata
es polimórfica y solo es predominante el patrón imitativo de la coral en las
zonas donde coincide con ella y sus zonas fronterizas, ya que en los demás lugares
este patrón no produce beneficios.

<> · Selección sexual <>:
WikipediA - 13/11/2017, 21:15, Lunes.;
La selección sexual es un concepto clave de la teoría de la
evolución acuñado por Charles Darwin en su libro El origen de las especies
para explicar el desarrollo de caracteres sexuales secundarios en los seres vivos
que parecían no responder a la selección natural, es decir, a la supervivencia del
mejor adaptado. En líneas generales, postula que ciertos rasgos presentes son el
resultado de la competencia entre individuos de un mismo sexo por el acceso a la
cópula (selección intrasexual) y de la selección por parte de uno de los sexos,
usualmente las hembras, de individuos del sexo opuesto (selección intersexual).
Se relaciona directamente a este mecanismo evolutivo con el dimorfismo
sexual y con la presencia de rasgos morfológicos exagerados. Muchos de estos rasgos
morfológicos llegan a suponer una merma en las capacidades adaptativas y de supervivencia
de los animales que los poseen. Para explicar su desarrollo, dentro de la selección
sexual, se postulan dos hipótesis denominadas «selección autorrefozante de Fisher»
y «del hándicap "¿de la desventaja?"», concebidas por Ronald Fisher y Amotz Zahavi
respectivamente. La primera de ellas explica este desarrollo exagerado de
estructuras fisiológicas en un sexo por la identificación directa de éstas con
genomas superiores por parte de sus parejas sexuales.La segunda de las hipótesis
indica que la presencia de rasgos morfológicos exagerados lastran la supervivencia
de los organismos que los manifiestan; sus parejas sexuales sentirían preferencia por
ellos por se capaces de sobrevivir a de la desventaja o hándicap que les
suponen estos rasgos.

(22/12/2017, 11:03, Viernes.)
Generalidades
El desarrollo del concepto de selección sexual fue una parte importante
de la obra de Charles Darwin El origen de las especies, publicada en 1859,
donde el autor le daba una gran importancia dentro de la selección natural. Darwin
explicaba mediante este mecanismo ciertos aspectos evolutivos tales como el desarrollo
exagerado de estructuras anatómicas o la presencia de colores llamativos que parecían
no ajustarse a la teoría de la selección natural donde los rasgos seleccionados para
prevalecer a lo largo del tiempo eran aquellos que aumentaran las probabilidades de
supervivencia. El naturalista británico conocía las ideas del anatomista John Hunter
sobre las diferencias entre caracteres sexuales primarios, relacionados con la
cópula, y los secundarios, relacionados con la atracción entre sexos. Basándose en
ello definió la selección sexual de tal modo que pudiera explicar la presencia de
estas características como resultado de una elección por parte de uno de los sexos
aunque no fue capaz de dar respuesta a por qué los individuos de este sexo mostraban
predisposición hacia ese rasgo. El término fue ampliado por Darwin en otro libro,
El origen del hombre y la selección en relación al sexo de 1871, ligándolo
además a la evolución de la especie humana como explicación para la existencia de
las diferentes razas.

La selección sexual fue reinterpretada en 1930 por el biólogo evolucionista
Ronald Aylmer Fisher. La nueva definición dada suponía que ciertos rasgos que en
la actualidad pueden parecer poco ventajosos para los organismos aparecieron
siendo lo contrario. Así, en un primer momento, la elección de estas características
tenía como único objetivo proveer a la descendencia de diversas ventajas en términos
de supervivencia. Esta mayor tasa de supervivencia supondría que gradualmente las
poblaciones naturales fueran diferenciándose por selección natural, un sexo a poseer
el rasgo y el otro por tener predisposición hacia él. Con el paso del tiempo,
ambos caracteres se vieron favorecidos al retroalimentarse uno de otro, llegando
a superar el objetivo inicial.
Es razonable pensar que, al igual que en el resto de especies animales,
la selección sexual haya actuado en la evolución del ser humano en mayor o menor
medida. Ya Darwin señaló un probable origen de la variedad racial humana en la
selección sexual aunque hoy día se considera que este proceso ocupó un papel muy
secundario debido a que los grupos humanos raramente se ha encontrado aislados
genéticamente y a que ha existido un alto mestizaje a lo largo de su evolución.
También apuntó a la selección sexual como responsable de la pérdida de vello
corporal de forma similar a lo ocurrido en otros primates. Diversas investigaciones
actuales relacionan las pautas humanas de elección de pareja, conductas y preferencias
físicas con la selección de caracteres que confieren ventajas genéticas a la
descendencia o con rasgos identificables con estados de salud de la pareja.
(22/12/2017, 15:40, Viernes.)
Inversión parental
La selección sexual únicamente puede ser comprendida teniendo presente
el concepto de inversión parental considerando este como el esfuerzo biológico
llevado a cabo por una pareja de una especie para concebir y criar a un descendiente.
El término fue ampliamente desarrollado por el investigador Robert Trivers,
basándose en los trabajos previos sobre gasto parental de Ronald Fisher, para
explicar la conducta sexual de especies animales.

En las especies con reprodución sexual heterogámica cada uno de los
parentales contribuye de forma no proporcionada a la creación de gametos. Por lo
general las hembras de estas especies producen una muy pequeña cantidad de óvulos,
esto se traduce en una cantidad limitada de descendientes ya que la disponibilidad
de óvulos se convierte precisamente en el recurso limitante de la reproducción. De
este modo la inversión parental de cada uno de los sexos es desigual, así como la
tasa potencial de reproducción, debido a que los costes de formación de los
gametos femeninos y el desarrollo de los embriones son muy altos para las hembras
en comparación con la producción de gametos masculinos. A esto habría que sumar que
las hembras deben también invertir gran cantidad de recursos en la crianza de sus
descendientes. Este esquema general es común en la mayoría de las especies animales
y principalmente en las poligínicas, donde cada macho puede potencialmente reproducirse
con varias hembras, mientras que los ejemplos de machos que realizan la mayor parte
de los cuidados parentales se da en las especies poliándricas, donde son las
hembras las que tienen acceso a varias parejas reproductoras. En las especies
monógamas es usual que ambos parentales compartan los esfuerzos en la cría
existiendo una cierta competencia entre sí para realizar el menor trabajo posible
aumentando el esfuerzo del otro.
De este modo suele aceptarse que el esfuerzo parental, es decir la
contribución conjunta de ambos sexos para el desarrollo de un descendiente, recae en
mayor medida en uno de los sexos, usualmente, la hembra. Al mismo tiempo el otro
sexo, por lo general el macho, contribuye principalmente en la etapa anterior
invirtiendo en el esfuerzo de apareamiento, esfuerzo para tener la posibilidad
de aparearse con las escasas hembras. En consecuencia suel admitirse que el parental
que invierte más en la reproducción será el responsable de la elección de su pareja
para asegurar su propia inversión mientras que el que invierte menos
debe competir por el acceso a la cópula.
Proporciones sexuales
Otro factor a tener en cuenta para comprender la selección sexual es la
proporción entre hembras y machos en las poblaciones naturales, el sex ratio,
(proporción de sexos) que suele se cercanoa 1:1 con igual número de descendientes de uno y otro sexo, y
resumido en el Principio de Fisher (aunque las diferencias en la mortalidad entre
machos y hembras modifica esta proporción con el paso del tiempo). La proporción de
sexos
influye decisivamente en la presión de la selección sexual en el sentido de que
establece la intensidad de la competencia por la cópula dependiendo de la cantidad
de parejas sexuales disponibles.

Resulta complicado comprender cómo la evolución no ha favorecido
proporciones diferentes considerando que la capacidad reproductiva de los machos
es superior a la de las hembras. Una cantidad mayor de hembras supondría mayor
número de individuos en la siguiente generación dado que cada macho podría fecundar
a varias de ellas. Diversos estudios han demostrado que un cambio en estas proporciones
es, al contrario de lo que podría pensarse, desfavorable para las poblaciones debido
a su inestabilidad a las mutaciones. Supongamos una especie en la que los machos
y las hembras estuvieran en proporción 1:3, el triple de hembras que de machos, y
cada apareamiento diera lugar a un único descendiente. En ella cada macho podría
tener al menos tres descendientes en la primera generación y cada hembra uno. En
esta situación una mutación cualquiera transmitida por un macho pasaría rápidamente
a la población reduciendo gradualmente su variabilidad genética.
Ronald Fisher propuso en 1930 que debían existir mecanismos que permitieran
a una población con las proporciones sexuales desplazadas respecto a su óptimo
evolucionar a través del tiempo hasta recuperarlas independientemente del sistema
de herencia del sexo que poseyera la especie, incluida la determinación del sexo
ligado a las condiciones físicas durante el desarrollo del embrión. Experimentos
desarrollados con Drosophila melanogaster han demostrado que efectivamente
existen tales mecanismos, aunque se desconoce su funcionamiento, mediante el estudio
de la evolución de la proproción de sexos en diferentes poblaciones. Igualmente
debían existir mecanismos mediante los cuales las especies en las que su proporción
de sexos óptimo fuese diferente a 1:1 lo mantuvieran a lo largo del tiempo.

Ejemplo de especies con sus proporciones desplazadas respecto de 1:1
puede ser las de aquellas afectadas por la competencia local por el apareamiento,
mecanismo que tiene lugar entre los mismos hermanos por las hembras. Desde el
punto de vista de los progenitores dado que sus descendientes varones van a competir
entre ellos mismos por las hembras y sólo una parte de ellos podrá reproducirse
resultará tanto más favorable concentrar los esfuerzos en producir hembras, todas
las cuales se reproducirán.
Los sesgos en las proporciones entre machos y hembras no solo tienen lugar
por la capacidad futura de los descendientes para aparearse sino también por la
capacidad de los mismos padres en generar hijos más competitivos. En el caso de
ciervo europeo, Cervus elaphus, la capacidad para copular con las hembras
está relacionada con la fuerza y salud física de los machos. Estos ciervos compiten
entre ellos mediante luchas y demostraciones físicas. La fuerza de estos machos dependerá
de de la alimentación y cuidados que sus padres hayan podido darles. Se sabe que
en esta especie existe un sesgo hacia la producción de machos cuando la hembra
que los produce es la hembra dominante del grupo. En este caso la hembra dominante
producirá machos mejor desarrollados y el balance de esfuerzos en su cría será
positivo al reproducirse estos con mayor probabilidad. Al mismo tiempo existe
un sesgo hacia la producción de hembras en los casos en que sus madres no sean
las dominantes, de este modo se aseguran la reproducción de sus descendientes.
Selección intrasexual frente a la selección intersexual
Las diferencias en la inversión parental entre progenitores y en las
proporciones sexuales tienen varias consecuencias. Por una parte, que el individuo
del sexo que más invierte en la reproducción deba asegurarse de seleccionar una
pareja sexual adecuada para optimizar su inversión. Por otra parte, que los individuos
pertenecientes al sexo mayoritario en número o en capacidad reproductora deban
competir entre sí por la cópula. La intensidad de la selección sexual es proporcional
al grado de competencia entre los individuos por los apareamientos. Estas premisas
son la base para la diferenciación de dos mecanismos dentro de la selección sexual
que suelen actuar
conjuntamente, la selección intrasexual y la selección intersexual. La selección
intrasexual es aquella que favorece el éxito reproductivo de un sexo por competencia
directa con individuos del mismo sexo, por ejemplo favoreciendo
rasgos que den mayor capacidad para luchar. La selección intersexual es aquella
que favorece los rasgos que aumentan el éxito reproductivo al aumentar la atracción
sobre el otro sexo.
Selección intrasexual
Dentro de la competencia entre individuos del mismo sexo por el acceso
a los del sexo contrario tienen especial importancia las luchas, bien sean reales
mediante el uso de estructuras corporales a modo de armas o bien sean rituales,
con la exposición ante el oponente de esas mismas armas pero sin llegar al combate.
Estos combates, por lo general llevados a cabo por los machos, pueden producirse
por el acceso a las hembras o por el acceso a los lugares donde estas se encuentran
y generalmente son más intensas conforme la hembra se va mostrando más receptiva.
Aunque son muy llamativos los ejemplos combativos llevados a cabo por muchos grandes
mamíferos, existen otras muchas variantes de esta competencia de manera menos
sangrienta.
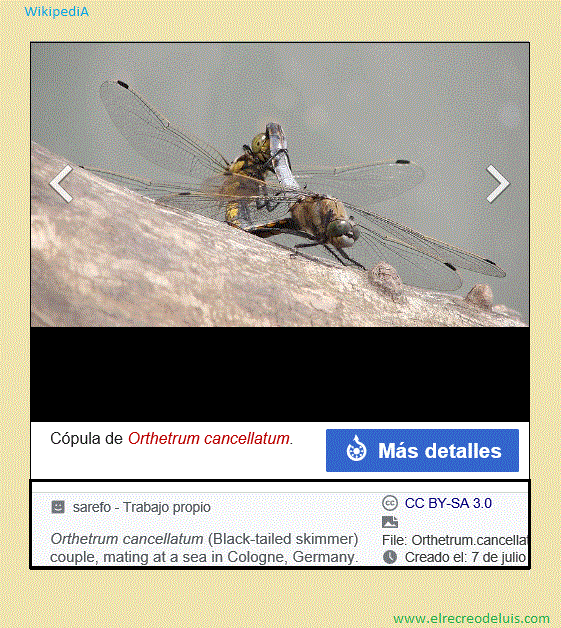
La competencia espermática es aquella en la que la propia cópula de un
macho actúa impidiendo que los gametos de otros machos fecunden a la hembra. La
competencia espermática parece ser la responsable de las marcadas diferencias
entre los genitales de especies muy cercanas filogenéticamente, especialmente en
insectos, y quizás por ello poseen un fuerte mecanismo de especiación. Existen
numerosos ejemplos de este tipo de competencia dentro del mundo de los insectos.
La libélula Orthetrum cancellatum, por ejemplo, tiene una particular forma
de reproducción donde la hembras se aparea con varios machos no siendo inmediantemente
fecundada por ninguno de ellos y guardando una espermateca en su interior. Los machos
de esta especie han desarrollado en su aparato reproductor una estructura preparada
para extraer del cuerpo de la hembra la espermateca de los machos que la han
fecundado con anterioridad. Otra especie similar, Crocethemus erythracea,
inyecta su espermateca con gran fuerza dentro del cuerpo de la hembra gracias a
un apéndice especial de su pene para situarla por delante de la de su competidores.
Existen otros ejemplos dentro de los invertebrados de especies que cierran
literalmente el orificio genital femenino como ocurre en el parásito acantocéfalo
Moniliformes dubius. La competencia por las hembras puede llegar a ser tal
que el macho de Xylocoris maculipennis, un hemíptero, copula con otros
machos inyectándoles en sus tejidos esperma que se traslada hasta las gónadas para
luego llegar a las hembras cuando ten lugar la cópula.
Un caso muy particular de selección sexual por competencia intrasexual
podría darse, de ser ciertas las consideraciones de ciertos autores, en las plantas
con flores. Aunque en líneas generales se acepta que la evolución de los caracteres
sexuales secundarios nada tiene que ver con factores de competencia y por tanto
con selección sexual cabe la posibilidad de que sí ocurra así con caracteres sexuales
primarios. Según ciertas teorías el desarrollo de las flores, su variada morfología,
colorido, posición tamaño de su estigma y el estilo podría responder a la necesidad
de las plantas que las poseen de atraer ponilinzadores. Caracteres como la presencia
de néctares y periantos llamativos pueden haber evolucionado a partir de la
competencia directa entre unos individuos y otros por aves e insectos que funcionaran
como vehículos para el polen.
Selección intersexual
Dado que para las hembras el coste de la reproducción es muy grande
en relación al de los machos, la elección de éstos es muy delicada para ellas. Los
criterios que pueden tener estas hembras son muy variados. Las hembras de algunas
especies deciden qué macho es más capaz atendiendo a la capacidad que tiene este
de suministrarles alimento o cobijo. Otras lo hacen en función del tamaño de
alguna parte de su cuerpo o de la complejidad de su ornamentación como ocurre con
las hembras de Triturus vulgaris y su preferencia por machos con grandes
crestas dorsales. Estos criterios tienen una base genética en tanto que supuestamente
la capacidad de los machos para la caza o para la elaboración de nidos estará
relacionada con la carga genética de éstos y esas mismas capacidades serán
trasmitidas a la prole.

La capacidad del parental para suministrar mejores recursos a la hembra
y a sus descendientes está estrechamente relacionada con la capacidad de supervivencia
que tendrá la descendencia. Se dan casos muy interesantes en aves donde se relacionan
las capacidades del macho para proporcionar alimentos a la hembra durante el
cortejo con la capacidad que tendrá después para suministrárselos a sus hijos.
Ocurre así con la golondrina de mar (Stema fuscata) donde muchas parejas
reproductivas en años anteriores se separan si la hembra considera que el macho ha
perdido esta capacidad para conseguir alimento un año concreto. Algunas ranas machos
son seleccionadas por su canto más potente y duradero, carácter que puede estar
revelando que el hábitat en el que viven -y en el que se sesarrollarán su crías-
es más rico ya que permite al macho una inversión tan importante.

Aunque es, de nuevo, en invertebrados donde se dan los más curiosos
ejemplos de este tipo de selección. La hembra de la mosca colgante, Hylobittacus
apicalis, seleccionará para el apareamiento al macho que le suministre como
alimento un insecto de mayor tamaño. Es más, este insecto permitirá que la cópula
sea más larga -y sean más los óvulos fecundados- en función del tamaño de la presa.
Gana la hembra, pues podrá transformar el alimento en huevos y gana el macho que se
reproduce. Pero no siempre se dan estas circunstancias, algunos machos de Hylobittacus
apicalis son capaces de forzar a la hembra y aparearse con ella sin haberles
llevado alimento. Esta situación es claramente desfavorable para la hembra que
deberá buscar alimento y favorable para el macho que ganará tiempo respecto a
otros congéneres para reproducirse.
Existen también experimentos que intentan demostrar la teoría de que
algunas hembras de algunas especies son capaces de elegir el macho con mejor
genética sin ningún tipo de demostración por parte de este, considerando que la
mejor genética es aquella que aumentea la capacidad de supervivencia de los
descendientes. En uno de estos experimentos tomaron varias hembras de Drosophila
melanogaster y se las dividió en dos grupos. Uno de ellos era forzado a aparearse
con un único macho tomado por los investigadores al azar mientras que al otro se les
mostraban a las hembras varios machos para que pudiera aparearse con el que consideraran
oportuno. Posteriormente las crías de ambos grupos fueron sometidas a la presión
de depredadores mostrando las de aquellas hembras que tuvieron libertad para
aparearse mejor capacidad de supervivencia...
(23/12/2017, 14:21, Sábado.)
...Teóricamente, y suponiendo que no hubiera existido algún tipo de competencia.
intraespecífica entre los machos, las hembras que pudieron elegir tomaron a los
machos mejor preparados y por ello sus descendientes eran capaces de sobrevivir a
la depredación.
La selección sexual y los rasgos morfológicos exagerados
El desarrollo en machos de elementos ornamentales muy elaborados como
caracteres sexuales secundarios es uno de los resultados de la selección sexual,
estudiado desde hace años como el ejemplo más representativo de esta. Muchas de estas
características se relacionan directamente con la competencia intraespecífica, tal
es el caso del tamaño de cuernos y astas utilizados en combates. En otras ocasiones
son claros ejemplos de selección por parte de las hembras de características
morfológicas en los machos, como la presencia de largas plumas o colores vistosos.
Muchos de estos adornos pueden parecer incluso desfavorables para la supervivencia
de los mismos individuos que las poseen y de su descendencia por disminuir su capacidad
de movimientos o de mimetismo. El caso paradigmático de adornos corporales sobredesarrollados
se presenta en el pavo real (Pavo cristatus) donde el macho exhibe en su cola unas
largas plumas con ocelos de color azul metálico. El estudio de la evolución de
este carácter fue parte primordial de los primeros estudios sobre selección sexual
llevados a cabo por Charles Darwin en El origen del hombre. Actualmente son
dos las teorías más importantes que se han propuesto para explicar el desarrollo
de ornamentos elaborados por selección de beneficios genéticos, la hipótesis de
Fisher y la hipótesis del hándicap (desventaja).

El buen gen. Fisherian runaway - Selección autorreforzante
La hipótesis de Fisher o Fisherian runaway, desarrollada por Ronald
Fisher, postula que los ornamentos elaborados de los machos son seleccionados
por las hembras, simplemente, porque son más atractivos para estas. Esta hipótesis,
también llamada del buen gen, postula que la atracción por un determinado
carácter estaría relacionada con que el rasgo en cuestión es la exteriorización de
un genotipo superior.
El tamaño de la cola de algunas aves, por ejemplo en la viuda de cola
larga (Euplectes progne), puede estar relacionado con la capacidad de vuelo
de estas. Las hembras en un principio seleccionarían al macho con la cola más
larga, mejor volador, y sus rasgos se transmitirían a su descendencia. De esta
manera se aseguraría la hembra tener descendientes con mejores capacidades para
el vuelo. El sistema se retroalimentaría, colas más largas, mayor éxito reproductivo,
descendientes con la cola más larga. En las hembras mientras tanto la existencia
de un gen que las predispusiera para seleccionar al macho con mayor cola también
se vería beneficiado.

Finalmente dado que hubo una selección en las hembras para aparearse con
machos de cola larga el rasgo más importante a trasmitirse dejará de ser el inicial,
la capacidad de vuelo, para transmitirse el rasgo de ser más atractivo ante las
hembras. Así, aunque el tamaño de la cola llegue a tal punto que comience a ser
un rasgo desfavorable para la especie, esto ya no tendrá importancia. Las hembras
de la viuda de cola larga, predispuestas ya a aparearse con machos atractivos,
elegirán a éstos para que sus descendientes sean más atractivos también ante otras
hembras independientemente de su capacidad de vuelo.
El principio de la desventaja (del hándicap)
Por su parte el principio del hándicap, desarrollada por Amotz Zahavi,
postula que algunas hembras seleccionan machos con ornamentos que suponen una
desventaja en la supervivencia precisamente por esto, porque son una desventaja,
un hándicap. En el mismo ejemplo anterior, un macho de ave con una cola muy
larga puede setar más expuesto a los depredadores que otro con la cola más corta.
Que este macho se presente ante la hembra demostraría que su genética superior
le ha permitido, a pesar del hándicap de su cola, sobrevivir en perfectas condiciones.
Este macho sería mejor, por tanto, que otro macho de cola corta que ha tenido más
fácil sobrevivir.

Se suele diferenciar dos tipos de desventajas (hándicaps), uno de ellos llamado
desventaja (hándicap) revelador muestra que el macho es fuerte ya que vive a pesar de la tara
en contraposición a los machos que no tienen el rasgo. El otro tipo, la desventaja
(el hándicap) condicional, muestra que el macho se encuentra fuerte ya que ha sido
capaz de desarrollar la característica en contraposición a quellos machos enfermos
o mal alimentados, que no han podido desarrollar el rasgo. Un ejemplo puntual de
este extremo es que la presencia de parásitos en aves tiene como principal síntoma
externo una menor pigmentación en las plumas. El autor propone explicaciones
basadas en la teoría de la desventaja (hándicap) para un gran número de situaciones de dimorfismo
sexual. Un de los casos estudiados es el de especies en las que los rasgos expresados
suponen una reducción en la visión. Ocurre en determinadas especies de primates,
como el orangután (Pongo pygmaeus) donde los amplios repliegues cutáneos presentes
alrededor de los ojos dificultan notablemente la capacidad de éstos para relacionarse
con su medio ambiente. Según las hipóteis de Zahavi la supervivencia de un macho
con la visión reducida pondría de manifiesto, ante las hembras, una superioridad
genética de este macho frente a otros que no tuvieran esa tara.
<> · selectivo, va <>:
R.A.E.:
1. adj. Que implica selección.
2. adj. Dicho de un aparato radiorreceptor: Que permite escoger una
onda de longitud determinada sin que perturben la recepción otras ondas muy
próximas.
·Semejante:
rae; 1. adj. Que semeja o se parece a alguien o algo. 4. adj. Geom. Dicho
de una figura: Que es distinta a otra solo por el tamaño y cuyas partes guardan todas
respectivamente la misma proporción. 5.m Semejanza, imitación. 6. m. prójimo.
<>·Semen<>:
rae; Del latín ·semen·. 1. m. Conjunto de espermatozoides y sustancias fluidas que se producen en el aparato genital masculino de los animales y de la especie human. 2. m. Bot. Semilla de los vegetales.
·Senescente: rae; Del latín ·senescens·, -entis.. 1. adj. Que empieza a envejecer.
<>·Seno<>:
rae; Del latín 'sinus'. 1. m. Concavidad o hueco. 2. m. Concavidad que forma una cosa encorvada. 3. m. pecho (// cada una de las mamas de la mujer). 4. m. Espacio o hueco que queda entre el vestido y el pecho. Sacó del seno una bolsa 5. m. Matriz de la mujer y de las hembras de los mamíferos. 6. m. Parte interna de algo. El seno del mar. El seno de una sociedad. 7. m. Parte de mar que se recoge entre dos puntas o cabos de tierra. 8. m. regazo (// cosa que recibe en sí a otra amparándola). 9. m. Anat. Cavida existente en el espesor de un hueso o formada por la reunión de varios huesos. El seno frontal. El seno maxilar. 10. m. Arq. Espacio comprendido entre los trasdoses de arcos o bóvedas contiguas. 11. m. Geogr. golfo (// porción de mar que se interna en la tierra). 12. m. Geom. Cociente entre el cateto opuesto a un ángulo y la hipotenusa. 13. m. Marina. Curvatura que hace cualquier vela o cuerda que no esté tirante. 14. Med. Pequeña cavidad que se forma en la llaga o postema.
<>Seno<>:
rae;
12. m. Geom. Cociente entre el cateto opuesto a un ángulo y la hipotenusa.
<>·Separar<>:
rae; Del latín ·separâre·. 1. tr. Establecer distancia, o aumentarla, entre algo oalguien y una persona, animal, lugar o cosa que se toman como punto de referencia. 2. tr. Formar grupos homogéneos de cosas que estaban mezcladas con otras. 3. tr. Considerar aisladamente cosas que estaban juntas o fundidas.
·Septo:
rae; Zoología; Tabique que divide de un modo completo o incompleto una cavidad
o partes del cuerpo en un animal.
·Ser:
rae; 1. m. Esencia o naturaleza. 3. m. Ser humano. Es un ser admirable.
/// Ser:
Elemento dotado de vida. Seres orgánicos. Seres vivos.
/// <>Ser vivo<>:
www.hiru.eus - (13/02/2017. Lunes. 15:17);
Un ser vivo u organismo
es un conjunto material de organización compleja, en la
que intervienen sistemas de comunicación molecular que lo relacionan internamente
y con el medio ambiente en un intercambio de materia y energía de una forma
ordenada, teniendo la capacidad de desempeñar las funciones básicas de la vida
que son la nutrición, la relación y la reproducción, de tal manera que los seres
vivos actúan y funcionan por sí mismos sin perder su nivel estructural hasta su
muerte.
La materia que compone los seres vivos está formada en un 95% por cuatro
elementos (bioelementos) que son el carbono, hidrógeno, oxígeno y nitrógeno, a
partir de los cuales se forman biomoléculas.
● Biomolécula orgánicas o principios inmediatos: glúcidos, lípidos,
proteínas y ácidos nucleicos.
● Biomléculas inorgánicas: agua, sales minerales y gases.
Estas moléculas se repiten constantemente en todos los seres vivos, por lo
que el origen de la vida procede de un antecesor común, pues sería muy improbable
que hayan aparecido independientemente dos seres vivos con las mismas moléculas
orgánicas. Se han encontrado estromatolitos con una antigüedad de 3700 millones
de años, por lo que la vida podría haber surgido sobre la Tierra hace 4100-3800
millones de años.
Todos los seres vivos están constituidos por células (véase teoría celular).
En el interior de éstas se realizan las ecuencias de reacciones qúmicas,
catalizadas por enzimas, necesarias pra la vida.
La ciencia que estudia los seres vivos es la biología.
<>·Serología<>:
R.A.E.; De sero- y -logía.
1. f. Estudio químico y bioquímico de los sueros, especialmente del
suero sanguíneo.
<>·Seta<>:
rae; De origen incierto. 1. f. Cualquier especie de hongo, comestible o no, con forma de sombrilla, sostenida por un pedicelo.
<>·Sexo<>:
rae; Del latín ·sexus·. 1. m. Condición orgánica, masculina o femenina, de los animales y las plantas. 2. m. Conjunto de seres pertenecientes a un mismo sexo. Sexo masculino, femenino. 3. m. Órganos sexuales. 4. m. Actividad sexual.
/// ·Sexual:
Wikipediá - REPRODUCCIÓN SEXUAL - (20/11/2016); La reproducción sexual es el
proceso de crear un nuevo organismo descendiente a partir de la combinación de material
genético de dos organismos de una misma especie, comenzando con un proceso que se
denomina meiosis, que es un tipo especializado de división celular; el cual se produce
en organismos eucariotas. Los gametos son los dos tipos de células especiales, uno
más grande, el femenino, y otro más pequeño, el masculino. La fusión de estas dos
células se llama fertilización y ésta crea un cigoto, que incluye material de los dos
gametos en proceso que se llama recombinación genética donde el material genético, el ADN,
se empareja para que las secuencias de los cromosomas homólogos se alineen. A continuación
se producen otras dos divisiones celulares más, pra producir cuatro células hijas
con la mitad de cromosomas de cada una de las dos células del padre, y el mismo número
que tienen los padres, aunque puede ocurrir la auto-fertilización. Por ejemplo, en la
reproducción humana cada célula humana contiene 46 cromosomas (23 pares), mientras que
los gametos, sólo contienen 23 cromosomas, así el hijo tendrá 23 cromosomas de cada
padre recombinadas genéticamente en 23 pares.
La evolución de la reprodución sexual es una de los principales rompecabezas debido
al hecho que la reproducción asexual la debe poder superar porque cada organismo joven
puede cargar sus propias descendientes, lo cual implica que la población asexual tiene
una capacidad intrinseca para crearse más rápido con cada generación. Este costo de
50% es una desventaja de aptitud de la reproducción sexual. Lo que se llama el costo
doble del sexo incluye este costo mientras el hecho que un organismo sólo puede pasar 50%
de sus propios genes a cada uno de sus descendientes representa un segundo costo. Una
ventaja definitiva de la reproducción sexual es que impide la acumulación de mutaciones
genéticas.
La selección sexual es un tipo de selección natural, en la cual algunos individuos
se reproducen con más éxito que otros en una población debido a la selección intersexual.
Se ha descrito como una 'fuerza evolutiva que no existe en las poblaciones asexuales'.
Las procariotas, cuyas células tienen material adicional o transformado, se reproducen
a través de la reproducción asexual aunque a veces, en la transferencia genética
horizontal, pueden exhibir procesos como la conjugación procariota, la transformación
y la transducción, las cuales son similares a la reproducción sexual a pesar de no
acabar en reproducción.
·Sifón:
rae; 1; Tubo encorvado que sirve para sacar líquidos del vaso que los contiene,
haciéndolos pasar por un punto superior a su nivel.
/// <>·Sifonado<>:
www.plantasyhongos.es - Universidad de Extremadura - (07/01/2017-Sábado-16:15); Sin paredes que separen a los nucleos en células.
·Sifonal:
Aplicase a un tipo de organización del talo caracterizado por presentar una gran
célula plurinucleada de forma laminada, tubular o filamentosa.
·Silvestre:
rae; Del latín ·silvestris·.1. adj. Dicho de una planta: Criada naturalmente
y sin cultivo. 2. adj. Dicho especialmente de un lugar: Agreste, inculto o no cultivado.
3. adj. Dicho de un animal: No domestido.
<> · Simbiogénesis <>:
WikipediA - 20/11/2017, 15:55, Lunes.
La simbiogénesis es el resultado de endosimbiosis estables a
largo plazo que desembocan en la transferencia de material genético, pasando
parte o el total del ADN de los simbiontes al genoma del individuo resultante.
Del proceso simbiogenético surge un nuevo organismo en el que en su célula o
células se encuentran integrados los simbiontes.
...
<>·Simbionte<>:
rae; 1. m. Biol. Individuo asociado en simbiosis.
·Simbiosis:
rae; Asociación de individuos animales o vegetales de diferentes especies,
sobre todo si los simbiontes sacan provecho de la vida común. /// También
hongos con plantas.
·Símbolo:
Rae; Del latín ·symbôlus, y este del griegos? ·µß????· sýmbolos ·. 1. m.
Elemento u objeto material que, por convención o asociación, se considera representativo
de una entidad, de una idea, de una cierta condición, etc. La bandera es símbolo
de la patria. La paloma es el símbolo de la paz. 3. m. Ling. Representación
gráfica invariable de un concepto de carácter científico o técnico, constituida
por una o más letras u otros signos no alfabetizables, que goza de difusión internacional,
y que, a diferencia de la abreviatura, no se escribe con punto pospuesto; por ejemplo,
N, He, Km y , por norte, helio, kilómetro y euro. respectivamente.
/// Símbolo químico:
WikipediA (09/04/2016); Los símbolos químicos son abreviaciones o signos que se
utilizan para identificar los elementos y compuestos químicos. Algunos elementos
de uso frecuente y sus símbolos son: carbon = C, oxígeno = O, nitrógeno = N,
hidrógeno = H, cloro = Cl, azufre = S, magnesio = Mg, aluminio = Al, cobre = Cu,
argón = Ar, oro = Au, hierro = Fe, plata = Ag, platino = Pt. Fueron propuestos en
1814 por Berzelius en remplazo de los símbolos alquímicos y los utilizados por Dalton
en 1808 para explicar su teoría atómica. Las mayoría de los símbolos químicos se
derivan de las letras griegas del nombre del elemento. principalmente en latín, pero
a veces en inglés, alemán, francés o ruso. La primera letra del símbolo se escribe
con mayúsculas, y la segunda (si la hay) con minúscula. Los símbolos de algunos
elementos conocidos desde la antigüedad, proceden normalmente de sus nombres en latín.
Por ejemplo, Cu de cuprum (cobre), Ag de argentum (plata), Au de aurum (oro) y Fe
de ferrum (hierro). Este conjunto de símbolos que denomina a los elementos químicos
es universal. Los símbolos de los elementos pueden ser utilizados como abreviaciones para
nombrar al elemento, pero también se utilizan en fórmulas y ecuaciones para indicar
una cantidad relativa fija del mismo. El símbolo suele representar un átomo del
elemento en una molécula u otra especie química. Sin embargo, los átomos tienen
unas masas fijas, denominadas masas atómicas relativas, por lo que también representa un mol.
·Simetría
Rae; Del latín ·symmetrîa·, y este del griego s?µµet??a symmetría.
1. f. Correspondencia exacta en forma, tamaño y posición de las partes de un todo.
2. f. Biol. Correspondencia que se puede distinguir, de manera ideal, en el cuerpo
de una planta o de un animal respecto a un centro, un eje o un plano, de acuerdo
con los cuales se disponen ordenadamente órganos o partes equivalentes. 3. f. Geom.
Correspondencia exacta en la disposición regular de las partes o puntos de un cuerpo
o figura con ralación a un centro, un eje o un plano.
·Simple:
rae; Del latín simple adverbio de simplus.. 1. adj. Constituido por un solo
elemento, no compuesto. 2. adj. Sencillo, sin complicaciones ni dificultades.
<>·Sinapomorfías<>:
WikipediA - 22/10/2017 - 17:43 - Domingo.;
En biología sistemática y, en particular, en cladística, una apomorfia
o apomorfía (griego "forma separada" es un rasgo o carácter biológico
evolutivamente novedoso, una novedad evolutiva derivada de otro rasgo perteneciente
a un taxón ancestral filogenéticamente próximo. El concepto de apomorfía se opone
al de plesiomorfia, que se refiere a los rasgos históricamente más antiguos, de
los cuales derivan las apomorfías.
La distinción entre apomorfia y plesiomorfaia no es absoluta, sino
relativa a un nivel determinado de la jerarquía sistemática:
▪ En el contexto de los mamíferos como subtaxón de los
tetrápodos, el martillo del oido es una apomorfia. En relación a este último,
el cartílago articular de los anfibios (una parte de la mandíbula inferior de
la que se deriva el martillo) es un carácter plesiomórfico.
▪ Si consideramos a los animales como un conjunto, la columna
vertebral es una apomorfia que caracteriza al subtaxón de los vertebrados. Pero si
queremos caracterizar la posesión de columna vertebral en un pájaro en relación
al taxón de los vertebrados, entonces el rasgo es plesiomórfico.
Sinapomorfías y autapomorfías
Cuando se somete a análisis un dendrograma, cladograma o filograma, las
apomorfías observadas (caracteres que difieren del ancestral) se pueden subdividir
en dos subgrupos:
▪ Sinapomorfías: aquellas apomorfías compartidas por
dos o más grupos.
▪ Autapomorfías: aquellas apomorfías que sólo aparecen
en un grupo.
El origen de novedades morfológicas es uno de los asuntos más debatidos
en biología evolutiva.
/// <>·Singamia<>:
TREA CIENCIAS - (04/12/2016-Domingo); Unión de dos células sexuales para dar un zigoto (fecundación).
<> · Sinopsis <>:
R.A.E.; Del latín tardio «synopsis», y este del griego
[σύνοψις "sýnopsis"], [συν-
"syn-" 'con-'] y [οψις "ópsis" 'vista'].
1. f. Disposición gráfica que muestra o representa cosas relacionadas
entre sí, facilitando su visión conjunta.
2. f. Exposición general de una materia o asunto, presentados en sus
líneas esenciales.
3. f. Sumario o resumen.
·Síntesis:
rae; Del latín synthesis, y este del gr sýnthesis. 1. f. Composición de un
todo por la reunión de sus partes. 2. f. Suma y compendio de una materia u otra cosa.
3. f. Quím. Proceso de obtención de un compuesto a partir de sustancias más sencillas.
/// ·Síntesis de ADN:
Buscar en el glosario, Replicación de ADN.
/// ·Síntesis de proteínas:
http:// proteinas.org.es/sintesis-proteinas. - (14/11/2016); Se conoce como síntesis
de proteínas al proceso por el cual se componen nuevas proteínas a partir de los
veinte aminoácidos esenciales. En este proceso, se transcribe el ADN en ARN. La sintesis
de proteinas se realiza en los ribosomas situados en el citoplasma celular.
En el proceso de síntesis, los aninoácidos son transportados por ARN de transferencia
correspondiente para cada aminoácido hasta el ARN mensajero donde se unen en la posición
adecuada para formar las nuevas proteínas.
Al finalizar la síntesis de una proteína se libera el ARN mensajero y puede volver
a ser leido, incluso antes de que la síntesis de una proteína termine, ya puede
comenzar la siguiente, por lo cual, el mismo ARN mensajero puede utilizarse por varios
ribosomas al mismo tiempo.
<> · Síntesis evolutiva moderna <>:
WikipediA - 16/11/2017, 15:49, Jueves.;
La síntesis evolutiva moderna (también llamada simplemente
nueva síntesis, síntesis moderna, síntesis evolutiva,
teoría sintética, síntesis neodarwinista o neodarwinismo
significa en general la integración de la teoría de la evolución de las especies
por la selección natural de Charles Darwin, la teoría genética de Gregor Mendel
como base de la herencia genética, la mutación aleatoria como duente de variación
y la genética de poblaciones. Los principales artífices de esta integración fueron
Ronald Fisher, J.B.S. Haldane y Sewall Wright.
Esencialmente, la síntesis moderna introdujo dos descubrimientos
importantes: la unidad de la evolución (los genes) con el mecanismo de la
evolución (la selección natural). También representa la unificación de varias
ramas de la biología que anteriormente tenían poco en común, especialmente la
genética, la citología, la sinstemática, la botánica y la paleontología.
...
·Sistema:
rae; Del latín tardio systêma y este del griego sýstema. 1. Conjunto de
reglas o principios sobre una materia racionalmente enlazados entre sí. 2. Conjunto
de cosas que relacionadas entre sí ordenadamente contribuyen a determinar un objeto
3. Bio. Conjunto de órganos que intervienen en alguna de las principales funciones
vegetativas. Sistema nervioso
<>·Sistema artificial de clasificación<>:
WikipediA - (14/10/2017, 14:15, Sabado.);
En Biología, un sistema artificial de clasificación es aquella organización
relacionada de elementos en el que la pertenencia de cada uno de ellos a diferentes
clases, depende de un criterio arbitrario adoptado por convención. Ejemplos de
tales criterios pueden ser la forma de desarrollo o el número de piezas florales.
Su ventaja es la de poseer un alto valor predictivo. El sistema artificial más
conocido fue, "Systema Naturae", donde se separan 23 clases de plantas con flores
de acuerdo con: la disposición de los sexos de las flores y el número, concrescencia,
inserción y relación de longitud de los estambres. Se añadia además una clase vigésima
cuarta de plantas sin flores (Cryptogamia) que incluía los helechos, musgos, algas,
hongos, etc... además de algunas plantas con flores difíciles de reconocer (
Ficus, Lemna). Así mismo incluyó los corales y las Porifera. Teofrasto,
considerado el padre de la Botánica, establecio una clasificación de 480 plantas en
árboles, arbustos, e hierbas. A su vez, las hierbas las subdividío en acuáticas y
terrestres, y agrupó a los árboles según la duración de sus hojas (perennifolios y
caducifolios).
<>·Sistema de determinación del sexo<>:
WikipediA - (15/10/2017, 13:14, Domingo.);
Un sistema de determinación del sexo es un sistema biológico que determina
el desarrollo de las características sexuales de un organismo (concretamente de
una especie). Se distinguen, generalmente, cuatro tipos de sistemas de determinación
del sexo:
● Determinación genética del sexo (DSG)=(GSD): aquella que está
determinada por factores contenidos en los cromosomas.
● Determinación cromosómica:
cuando existen cromosomas heteromórficos.
● Determinación génica:
cuando los cromosomas son homomórficos.
● Determinación haplodiploidía:
cuando uno de los sexos posee la mitad de la dotación cromosómica del otro sexo.
● Determinación ambiental del sexo (DSA)=(ESD): aquella que está
determinada por factores ambientales.
● Determinación por temperatura (DST)=(TSD):
establecido por la temperatura de incubación.
...
...
...
Sistema de determinación del sexo en plantas.
Hay que distinguir distintos tipos de plantas.
Dioicas: Cuando producen un único tipo de gametos. Por tanto,
una especie dioica es aquella que tiene plantas que sólo forman flores femeninas
o flores masculinas.
Monoicas: Son aquellas que en la misma planta se dan órganos
sexuales masculinos y femeninos. En concreto son monoicas las especies cuyas
plantas tienen ambos sexos pero en flores unisexuales separadas.
Si los sexos coinciden en la misma flor, entonces se denominan hermafroditas.
El determinismo genético del sexo en las plantas puede ser debido a
simples genes o estar relacionado con la presencia de cromosomas sexuales.
Un ejemplo del primer caso, determinación del sexo por genes, se de en
Ecbalium elaterium (pepinillo de los montones, que en las poblaciones del
norte de España se presenta como forma monoica y en el sur como dioica. Galán
propuso para esta especie un determinismo que consiste en una serie alélica, dando
lugar a plantas monoicas con una determinada combinación y dioicas cuando es otra.
En el caso de que sea una determinación por cromosomas sexuales, ha sido estudiada
en una hepática Sphaerocarpus donnelli, donde Allen observó diferencias
cromosómicas entre los gametofitos masculinos y femeninos. Aunque ambos tenían
el mismo número de cromosomas, en el femenino había un cromosoma X grande o en el
masculino un Y pequeño. También han podido ser identificados los cromosomas sexuales
de algunas Briofitas.
Abreviatura de uso común.
SDS = Sistema de determinación sexual
DSG = Determinación sexual genotípica. (En inglés, GSD = Genotypic sex determination).
DST = Determinación sexual por temperatura. (En inglés, TSD = Temperature-dependent sex determination).
DSA = Determinación sexual ambiental. (En inglés, ESD = Environmental sex determination).
/// Sistema endomembranoso: El sistema endomembranosos es el sistema de membranas internas de las células eucariotas que divide la célula en compartimentos funcionales y estructurales denominados orgánulos. Las procariotas no tienen un sistema endomembranoso y así carecen de la mayoría de los orgánulos
<>·Sistema natural de clasificación<>:
WikipediA - 15/10/2017 - 19:28 - Domingo. ;
Un sistema natural es aquella organización relacionada de elementos
que surge como una propiedad de la naturaleza. El concepto de sistema natural se
opone al de sistema artificial, en el que la pertenencia de los elementos a las
respectivas clases depende de un criterio artificial adoptado por convención. Por
el contrario, en el sistema natural deben estar contenidos datos específicos, que
son de vital importancia para dar un estudio más detallado acerca de la clasificación
de los seres vivos.
Nominalismo y Esencialismo
Una taxonomia puede ser considerada como un sistema natural desde el punto
de vista tanto del nominalismo como del esencialismo. Desde la perspectiva nominalista,
una clasificación, de los seres vivos es natural en tanto en cuanto refleja el patrón
de similitudes que observamos en la naturaleza. La "naturalidad" del sistema se
atribuye, por lo tanto, a la percepción humana, no a la naturaleza misma. Para el
esencialismo, una clasificación es natural porque revela grupos naturales reales,
no una mera coincidencia de semejanzas.
Historia del concepto de Sistema Natural
En biología, la creencia en el carácter natural de las clasificaciones
taxonómicas no estuvo siempre ligada a la creencia en una teoría que las explicase:
muchos naturalistas creían en la realidad de los taxones y el correspondiente ajuste
de las categorías taxonómicas, pero admitían el desconocimiento de la causa
responsable de tal ajuste.
A lo largo del siglo XIX, la teología natural ligó la creencia en el
Sistema Natural a la inmutabilidad de las ideas divinas. Fueron varios los
sistemas naturales (y varios también los criterios de naturalidad) propuestos a lo
largo de este siglo. En la inglaterra de 1820, por ejemplo, se manejaban cuatro
posibles sistemas naturales: el sistema de clasificación de Cuvier, el esquema
lineal y progresivo de Lamarck, el sistema dicotómico de Jeremy Bentham (1817) y
el sistema quinario o circular de William Sharp MacLeay.
Con el triunfo del evolucionismo, la naturalidad de las clasificaciones
se hizo radicar en la ascendencia común. De este modo, el sistema natural se
transformó en árbol filogenético.
<>·Sistemática<>:
WikipediA - 22/10/2017 - 18:34 - Domingo.;
La sistemática es un área de la biología encargada de clasificar
a las especies a partir de su historia evolutiva (filogenia). Se ocupa de la
diversidad biológica en un plano descriptivo y en uno interpretativo.
La unidad fundamental de la que parte toda la clasificación es la especie,
aquella agrupación de seres en la que todos sus miembros son capaces
de aparearse entre sí y producir descendencia viable y fértil. Para la designación
de las especies se utiliza la nomenclatura binomial de Linneo, consta de dos palabras
escritas en minúsculas y cursiva, la primera de ellas corresponderá siempre al género
y comienza con mayúscula y la segunda escrita en su totalidad en minúscula corresponde
a la especie. Los nombres de géneros y especies siempre van en una tipografía
diferente que el resto del texto, normalmente en itálicas para textos tipografiado
y subrayados en el caso de un texto escrito a mano. Los biólogos y paleontólogos
suelen incluir detrás del nombre completo de la especie el nombre de la persona
que describió, así como el año de publicación.
En los años cincuenta el entomólogo alemán Wili Hennig (fundador de la
cladística), sugirió un sistema de agrupación basado en la historia
evolutiva de los seres vivos (filogénesis), en el que tras estudiar todas las
características de un grupo y excluir aquellas que no marcan su desarrollo, define
una nueva agrupación, llamado clado, es necesario advertir que los rasgos
más obvios y acusados no necesariamente marcan el desarrollo evolutivo de un linaje.
Esta agrupación en clados presta más atención a los puntos en que los diversos clados
se bifurcan o separan entre sí, que a la agrupación por características morfológicas,
que en algunos casos, podrían ser secundarias y desvirtuar la lógica en el tiempo
del esquema arbóreo.
Origen
Desde que la vida surgió en sus formas más sencillas, hace aproximadamente
3.500 millones de años, los seres vivos se han ido diferenciando generación tras
generación en un proceso de evolución continua. Se estima que están descritas
alrededor de 2 millones de especies vivientes de animales y se infiere que deben
existir varios millones más. Si pensamos en el número de especies que surgen y las
extintas desde que apareció la vida, el número total asciende extraordinariamente.
La sistemática es un área de la biología que se desarrollo primero dentro
del campo de la historia natural, abarcando la parte descriptiva y sistemática de
la geología. Su método se centró inicialmente en la observación sistemática. La
rápida acumulación de datos descriptivos condujo a mediados del siglo XVIII a la
revolución linneana, con el establecimiento de convenios precisos de nomenclatura
y clasificación de los seres vivos, válidos con independencia de la nacionalidad,
la lengua o los objetivos precisos del investigador. Así que el sistema más
utilizado con algunas modificaciones es el "Linneano", creado por el botánico sueco
Carlos Linneo (1707-1778) en el siglo XVIII.
Así como otras ciencias que se ocupan de sistemas complejos y determinados
por una historia concreta, la sistemática se benefició del desarrollo de la
estadística descriptiva, desde finales del s.XIX y muy especialmente de la
estadística multivariante, a mediados del s. XX.
La diversidad de características que despliega la naturaleza viviente,
las semejanzas y diferencias, son el resultado de la evolución y dos causas generales:
1) Las relacionadas con la herencia. Todas las modificaciones, por muy
drásticas que sean, como las extremidades de los vertebrados, el ala de los murcielagos
o la aleta del delfín, parten de una misma estructura esquelética, que se altera
sólo dentro de límites amplios pero insalvables.
2) Las relacionadas con la adaptación. La evolución conduce a los caracteres
hacia grados mayores de adecuación para la función que los explica. Es la existencia
de la adaptación, uno de los efectos de la selección natural, la que obliga a la
biología a recurrir a fórmulas teleológicas en sus explicaciones.
Taxonomía y sistemática
Para el estudio de la clasificación de los organismos surgió una ciencia
llamada taxonomía (de la raíz griega taxis que significa 'ordenación').
La organización que establece la taxonomía tiene una estructura arbórea en la que las
ramas a su vez se dividen en otras y éstas se subdividen en otras menores. A cada
una de las ramas, ya sean grandes o pequeñas, desde su nacimiento hasta el final,
incluyendo todas sus ramificaciones, se les denomina taxón.
La taxonomía tiene por objeto agrupar a los seres vivos que presenten
semejanzas entre sí y que muestren diferencias con otros seres, estas unidades se
clasifican principalmente en siete categorías jerárquicas que son, por orden decreciente
de sus niveles:
Dominio - Reino - Phylum o División (Tipo) - Clase - Orden - Familia -
Género - Especie.
Estos ocho niveles a veces no son suficientes para clasificar de forma
clara a todos los seres vivos y es necesario en algunas ramas crear subdivisiones
intermedias, como suspendedor, suborden y super familias.
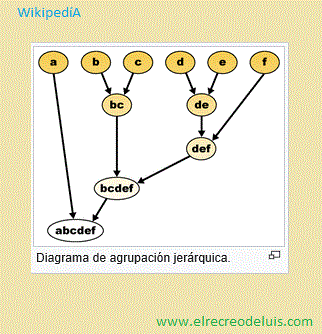
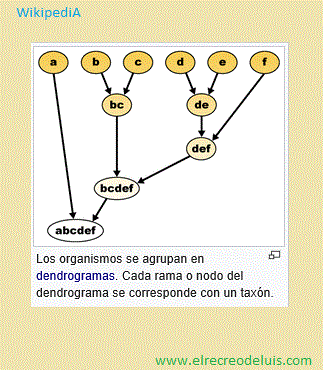
Herramientas taxonómicas
Dentro de las herramientas taxonómicas diseñadas para facilitar la
determinación de los organismos se encuentran las claves, éstas pueden ser
dicotómicas, politómicas o con sangría. Las dicotómicas tienen dos opciones, cada
una contiene una serie de caracteres contrastantes entre sí, está diseñada de tal
forma en que sólo una opción puede aceptarse, a cada par de alternativas a escoger
se la llama copla o dilema.
El objetivo de la clasificación es ordenar o disponer por clases a los
organismos. En la década de los ochenta se utilizaba una serie de palabras para
presentar un arreglo particular de acuerdo con algún principio de relación que
se creía existía en los organismos. Pero ya para 1990 comenzó a emplearse un
sistema de palabras y símbolos que denotaban conceptos.
En términos de clasificación biológica existen jerarquías, se refieren
a los rangos que denotan grupos ordenados en una secuencia de conjuntos
y subconjuntos, incluidos sucesivamente.
La clasificación artificial, agrupa a los organismos por atributos
designados de acuerdo a sus similitudes o sobre la base de su utilidad, en el
caso de seres vivos se hace sin que exista ninguna hipótesis de relación o
teoría biológica como ancestralidad común.
A diferencia de la clasificación artificial, la clasificación natural se
basa en las relaciones genealógicas y que sólo contienen grupos monofiléticos, los
cuales están conformados por grupos de especies que incluyen un ancestro y sus
descendientes. Desde la época de Linneo los taxónomos han intentado desarrollar
clasificaciones naturales, sin embargo hasta antes de la publicación de El origen
de las especies de Darwin, las clasificaciones no reflejaban la historia evolutiva
de los grupos.
De los conceptos aceptados en el siglo XXI se asume que la clasificación
natural debe reflejar relaciones filogenéticas entre las especies a clasificar.
La clasificación filogenética está basada en relaciones genealógicas
entre grupos de organismos.
Escuelas sistemáticas
Tres son las principales escuelas sistemáticas:
▪ Sistemática evolutiva. Encabezada principalmente por
J. Huxley, G. G. Simpson y E. Mayr, planteó por primera vez de un modo formal la
manera de reconstruir filogenias y de representarlas en forma de clasificaciones.
La sistemática evolutiva utiliza cuatro criterios principales: la discrepancia
morfológica, el nicho adaptativo, la riqueza en especies y la monofilia mínima.
Para clasificar, además de la filogenia, los evolucionistas se valen de criterios
como el hueco morfológico, el grado y la radiación adaptativa.
▪ Taxonomía fenética. Simultáneamente surgio la escuela
de R. R. Sokal y P. H. A. Sneath, que considera que la filogenia no puede conocerse
de manera objetiva; por tanto, su finalidad no es la de reconstruir filogneias sino
la de establecer clasificaciones estables. Se sustenta en técnicas matemáticas que
permiten establecer clasificaciones basadas en fenogramas, estos últimos armados
según el grado de similitud. La escuela fenética toma el máximo número de caracteres
disponibles sin preguntarse por su significado evolutivo, afirmando que no hay
forma de decidir si un carácter es una homología o una homoplasia.
▪ Sistemática cladista (Cladística). El entomólogo
alemán Wili Hennig, en 1950, propuso un método para determinar árboles filogenéticos
o cladogramas para convertirlos en clasificaciones. Sus ideas han sido desarrolladas
y seguidas por numerosos autores, sobre todo después de que su libro fuera traducido
del alemán al inglés en 1965, probablemente con mayoría entre los sistemáticos en
el último cuarto del siglo XX. La idea central es la monofilia; según los cladistas,
un grupo es monofilético si comprende la especie encestral de este grupo y a todos
sus descendientes. El criterio de reconocimiento de un grupo monofilético es la
identificación de al menos un carácter apomorfo compartido por todos los miembros
del grupo y heredado de su especie ancestral. La cladística actual utiliza el
análisis filogenético y el principio de parsimonia para elaborar cladogramas.
Cladogramas
La filogenia da continuidad a las relaciones de parentesco entre
entidades biológicas, sean éstas géneros o especies y generalmente se representan
por un diagrama ramificado llamado cladograma.
Los cladogramas son dendogramas construidos con base en las
relaciones cladísticas o de ancestralidad común de los taxones terminales, reconocidas
por caracteres evolucionados compartidos o sinapomorfias.
La topología es la forma geométrica del cladograma o la secuencia
de abyacencia absoluta de los taxones. Mientras que la raíz es el punto donde se
le da la dirección al árbol, aunque también se conoce como raíz al taxón empleando
para ubicar el punto de orientación de árbol. El enraizamiento es el
procedimiento mediante el cual se emplea al grupo externo para ubicar la raíz.
Disciplinas del campo sistemático
La taxonomía y la nomenclatura son disciplinas instrumentales, que son
más cercanas al concepto de técnicas que al de ciencias.
La larga lista de especialidades tiene que ver con la diversidad misma
que la sistemática investiga. La división inicial en Zoología y Botánica se fue
ampliando para sumar a la Microbiología y a la Virología. Cada Taxón mayor es el
centro de una especialidad, y sus grupos subordinados los son a su vez de otra.
Por ejemplo, la Dipterología que estudia a las moscas y mosquitos es parte de la
Entomología, que estudia a los insectos en general, que por su parte está subordinada
a la Zoología.
<>·Sistemática evolutiva<>:
WikipediA - 20/10/2017, 15:38, Viernes.;
La sistemática evolutiva o evolucionista es una escuela
de clasificación que tiene como finalidad determinar no solo la genealogía, sino
también la evolución de los diferentes linajes que componen un taxón.
Historia
Hasta los años 40 del siglo XX, la sistemática permaneció ajena a los
nuevos marcos teóricos aparecidos en la biología. Por supuesto, se había intentado,
desde Darwin y Haeckel, aplicar la teoría de la evolución a la sistemática, pero
con poco éxito. La mayor parte de los trabajos sistemáticos se limitaban a la mera
descripción; se describía las especies, se proporcionaban los medios para reconocerlas
y se situaban en la jerarqua linneana.
En la década de los 40, se produjo una revolución conceptual en la
sistemática; los sistemáticos se empezaron a interesar, no solo por la descripción,
sino por el esclarecimiento de la historia y la evolución de los seres vivos. Fruto
de estas nuevas inquietudes surgieron tres grandes escuelas sistemáticas: sistemáticas
evolutiva, sistemática fenética (o taxonomía numérica) y sistemática filogenética
(o cladística).
La Nueva Sistemática Evolutiva
La Sistemática Evolutiva fue fundada principalmente por J. Huxley,
E. Mayr y G.G.Simpson y planteó por primera vez de un modo formal la manera de
reconstruir filogenias y de representarlas en forma de clasificaciones.
El punto de partida de esta escuela es un volumen colectivo editado por
J.S.Huxley en 1940 titulado The New Systematics. La idea de base era que las
especies, que eran el objeto principal de los sistemáticos anteriores, están
formadas de poblaciones y que el estudio detallado de dichas poblaciones y de sus
variaciones sería particularmente fértil para discernir los procesos evolutivos.
La Nueva Sistemática (como se denominó al principio esta escuela) instaba,
pues, a centrar la investigación sobre las problaciones más que sobre las especies.

Para E. Mayr, ornitólogo que desarrolló las ideas de Huxley, era básico
que la Nueva Sistemática tomara en consideración los datos proporcionados por
disciplinas que entonces estaban en pleno desarrollo, como la citología, la genética,
la ecología, la fisiología o la etología, lo cual serviria para contrastar los
resultados obtenidos por la morfología, y a prestar una máxima atención a los
procesos a expensas de las estructuras.
Otra importante contribución al nuevo planteamiento de la sistemática
fue la del paleontólogo G.G.Simpson, según este autor.
▪ La sistemática tiene como unidad básica la población.
▪ La variación es un elemento esencial de la naturaleza y
de la definición de las poblaciones.
▪ Las poblaciones son sistemas dinámicos que evolucionan.
▪ La especie solo puede definirse en términos de dinámica,
de evolución, de genética, de relaciones dentro y entre las poblaciones y no en
términos de estructuras morfológicas fijas.
Métodos de la Sistemática Evolutiva
La sistemática evolutiva se fundamenta en la genealogía y en la similitud.
Otorga una importancia capital al grado de adaptación para establecer las
clasificaciones. El grado de adaptación es una unidad anagenética, es decir, un
estado de progreso, de perfección evolutiva.
Para evaluar este nivel de adaptación, la sistemática evolutiva utiliza
cuatro criterios principales:
▪ Discontinuidad morfológica. Si dos taxones son muy
diferentes se clasifican en grupos diferentes.
▪ Nicho adaptativo Una zona adaptativa excepcional
justifica un rango jerárquico más elevado.
▪ Riquezas de especies. Un grupo que contenga muchas
especies tendrá un rango más elevado que uno pobre en especies.
▪ Monofilia mínima Los grupos deben descender de un
mismo ancestro, pero no contener forzosamente todos los descendientes del mismo.
Los árboles evolutivos obtenidos aplicando estos criterios reciben el
nombre de filogramas.
Según Mayr, dados dos grupos hermanos, uno puede estar biológicamente
muy cercano a la especie ancestral (como los cocodrilos, en el cladograma adjunto),
mientras que el otro puede haber adquirido un nuevo nicho adaptativo y haber
evolucionado hacia un nuevo tipo (las aves en el cladogrma). en Estas condiciones,
si sus caracteres biológicos se han hecho muy diferentes, estos grupos tendrán
un rango diferente en la jerarquía linneana. Para evaluar el grado de diferencia
entre grupos hermanos, los evolucionistas se fundamentan en el número de autapomorfías,
es decir, de novedades evolutivas exclusias que determinan el taxón más evolucionado.
Así, las relaciones entre las aves y los reptiles actuales son las mostradas en el
cladograma siguiente. Según la sistemática evolutiva, dado que las aves han
acumulado numerosas autapomorfías, han ocupado un nicho adaptativo totalmente
diferente al de los reptiles actuales y poseen muchas más especies que ellos,
es justificado separarlas en un taxón propio; el taxón reptiles es monofilético
según la sistemática evolutiva (monofilia mínima), pero parafilético según la
cladística, ya que no incluye a todos los descendientes del antepadado común
de aves y reptiles.
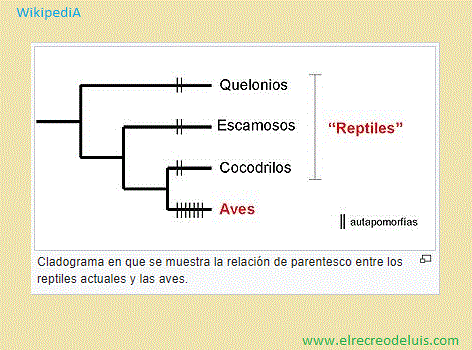
La sistemática evolutiva y la cladística se libraron durante casi
veinte años a una controversia en la que el dogmatismo no estuvieron siempre
ausentes. No obstante, después de algunos años, un gran consenso se estableció
entre los partidarios de la sistemática evolutiva para aceptar el principio
fundamental de la sistemática cladística: que los grupos de organismos no
pueden establecerse más que sobre la base de caracteres comunes y exclusivos,
las sinapomorfías, que puede pensarse que fueron heredados de un ancestro común.
<>·Sistemático, ca<>:
rae; Del latín tardío «systematîcus» y este del griego
'συστηματικος'
«systématikós».
1. adj. Que sigue o se ajusta a un sistema.
2. adj. Dicho de una persona: Que procede por principios, y con rigidez en
su tenor de vida o en sus escritos, opiniones, etc.
3. F.Biol. Estudio de la clasificación de las especies con arreglo a su
historia evolutiva o filogenia.
·Sol:
Rae; Del latín·sol·, ·solis·. Escrito con mayúscula inicial en acepción 1 en
contextos astronómicos. 1. m. Estrella luminosa, centro de nuestro sistema
planetario. El solsiticio es la época en que el Sol se halla en uno de los
trópicos. 2. m. Cualquier estrella luminosa que es centro de un sistema planetario.
3. m. Luz, calor o influjo del sol. Sentarse al sol. Tomar el sol. Entrar el sol
en una habitación. 4. m. Tiempo que el Sol emplea en dar aparentemente un vuelta
alrededor de la Tierra. 5. m. Unidad monetaria del Perú. 6. m. Alq. oro
(// elemento químico).
·Solubilidad:
Rae; 1. Cualidad de soluble.
/// <>Solubilidad<>:
WikipediA (19/02/2017 - Domingo. 20:42);
Solubilidad
es una medida de capacidad de disolverse de una
determinada sustancia (soluto) en un determinado medio (disolvente). Implícitamente
se corresponde con la máxima cantidad de soluto que se puede disolver en una
cantidad determinada de disolvente, a determinadas condiciones de temperatura, e
incluso presión (en caso de un soluto gaseodos). Puede expresarse en unidades de
concentració: molaridad, fracción molar, etc.
Si en una disolución no se puede disolver más soluto decimos que la disolución
está saturada. En algunas condiciones la solubilidad se puede sobrepasar
de es máximo y pasan a denominarse como soluciones sobresaturadas. Por
ejemplo, en el agua, se disuelve el alcohol y la sal, en tanto que el aceite y la
gasolina no se sisuelven. En la solubilidad, el carácter polar o apolar de la
sustancia influye mucho, ya que, debido a este carácter, la sustancia será más
o menos soluble; por ejemplo, los compuestos con más de un grupo funcional
presentan gran polaridad por lo que no son solubles en éter etílico.
Entonces para que un compuesto sea soluble en éter etílico ha de tener escasa
polaridad; es decir, tal compuesto no ha de tener más de un grupo polar. Los
compuestos con menor solubilidad son los que presentan menor reactividad, como
son: las parafinas, compuestos aromáticos y los derivados halogenados.
El término solubilidad se utiliza tanto para designar al fenómeno cualitativo
del proceso de disolución como para expresar cuantitativamente la concentración
de las soluciones. La solubilidad de una sustancia depende de la naturaleza del
disolvente y del soluto, así como de la temperatura y la presión del sistema, es
decir, de la tendencia del sistema a alcanzar el valor máximo de entropía. Al
proceso de interacción entre las moléculas del disolvente y las partículas del
soluto para formar agregados se le llama solvatación y si el solvente es agua,
hidratación.
·Soluble:
Rae; Del latín solubîlis. 1. adj. Que se puede disolver. 2. adj. que se
puede resolver. Problema soluble.
/// <>Soluble en disolventes apolares<>:
brainly.lat (19/02/2017 - Domingo. 23:51);
¿Por qué la grasa es soluble en disolventes orgánicos y no en el agua?
Primero que todo una grasa es un compuesto
organico que puede ser apolar, ahora bien, se sabe que un solvente polar disuelve
a un sustrato o soluto polar y un solvente apolar solo disuelve a un soluto apolar.
Entonces, el agua es un solvente polar, y la grasa es apolar, así que no podrá
disolverla. Sin embargo, el Eter Dietilico seria un buen solvente apolar para
disolver una grasa (de caracter analogo apolar).
·Soma:
rae; Del gr. s?µa sôma 'cuerpo'. 1. m. Biol. Totalidad de la materia corporal
de un organismo vivo, excepto los gametos.
<>·Sombrilla<>:
rae; Del diminutivo de ·sombra·. 1. f. Quitasol. = 1. Especie de paraguas o sombrilla usado para resguardarse del sol. 2. f. Costa Rica, Cuba, Mexico, El Salvador. paraguas (//utensilio para resguardarse de la lluvia).
<>·Soldadura<>:
R.A.E.;
1. f. Acción y efecto de soldar.
2. f. Material que sirve y está preparado para soldar.
3. f. Enmienda o corrección de algo. Este desacierto si tiene soldadura.
·Sub-:
rae; Del latín ·sub-·. 1. pref. Significa 'bajo' o 'debajo de'. Subsuelo,
subacuático. 2. pref. Indica inferioridad, subordinación, acción secundaria.
Subempleo, subdelegado, subarrendar.
·Subatómico, ca:
Rae; 1. adj. Fís y Quím. Dicho de un nivel estructural: Inferior al del
átomo. Los protones y los neutrones son partículas subatómicas.
<> · Subespecie <>:
R.A.E.:
1. f. Cada uno de los grupos en que se subdivide una especie.
<> · Subespecie <>:
WikipediA:
Se denomina subespecie a cada uno de los grupos en que se dividen
las especies, y que se componen de individuos que, además de los caracteres propios
de la misma, tienen en común otros caracteres morfológicos por los cuales se asemejan
entre sí y se distinguen de los de las demás subespecies. Desde el punto de vista
estrictamente sistemático o de la taxonomía, es la categoria taxonómica comprendida
entre la especie y la raza.
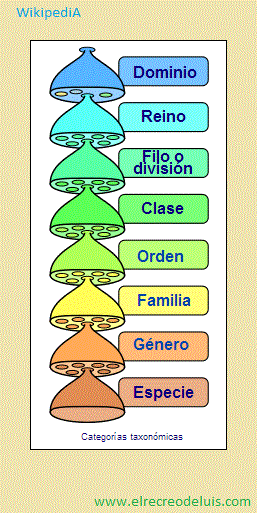
La subespecie es una parte de la especie formada por una o más poblaciones,
poco diferenciadas y con una distribución regional concreta. Esta definición
incluye las razas geográficas de muchos autores ('Rassenkreis'). Sin embargo, no
está muy clara la línea de separación entre los conceptos de raza y subespecie,
pues ambos taxones mantienen las características propias de la especie, es decir,
que parejas de individuos de distintas subespecies o razas (pero de la misma especie)
se pueden emparentar y producir descendencia viable y fértil. Además, solo se suelen
diferenciar en caracteres externos menores aunque reconocibles y regularmente
presentes. El término raza se utiliza con más frecuencia furea del ámbito científico,
por lo que, en taxonomía, solo se emplea (por tradición) para la distinción morfológica
en un número muy limitado de especies, como los animales domésticos. En el caso del
ser humano (Homo sapiens), la mayoría de los especialistas considera
inadecuado el uso de término raza para referirse a cada uno de los diversos o
diferentes grupos humanos.
Nomenclatura
La subespecie tiene una nomenclatura trinomial, es decir, formada por tres
nombres: el genérico, el específico y el subespecífico. Así, el perro tiene, como
nombre científico, el trinomen Canis lupus familiaris, donde Canis
es el género, lupus es la especie, y familiaris es la subespecie.
<>Subíndice<>:
rae;
1. m. Letra o número que se coloca en la parte inferior derecha de un
símbolo o de una palabra para distinguirlos de otros semejantes.
·Subseguir:
rae; 1. intr. Dicho de una cosa: Seguir inmediatamente a otra.
·Sudar: rae; Del latín ·sudâre·. 1. intr. Secretar sudor. 2. intr. Dicho de un árbol, de una planta o de un fruto: Destilar algunas gotas de su jugo. Sudar las castañas, o el café, después de tostados. 3. intr. Dicho de una cosa impregnada de humedad: Destilar agua a través de sus poros. Suda la pared, un botijo.
·Sudor:
rae; Del latín ·sudor·, ·-ôris·. 1. m. Líquido claro y transparente que segregan
las glándulas sudoríparas de la piel de los mamíferos y cuya composición química es
parecida a la de la orina. 2. m. Jugo que sudan las plantas. 3. m. Gostas que salen
y se destilan de las peñas u otras cosas que contienen humedad.
<>·Sugerir<>:
rae; Del latín ·suggerêre·. 1. tr. Proponer o aconsejar algo. Le sugerí que no trabajara tanto. 2. tr. Evocar algo o hacer pensar en ello. Esa canción me sugiere recuerdos de la infancia.
<>Suma<>:
rae; Del latín ·summa·.
1. Acción y efecto de sumar. 2. f. Agregado de muchas cosas, y más comúnmente
de dinero. 3. f. Lo más sustancial e importante de algo. 4. f. Recopilación de
todas las partes de una ciencia o facultad. 5. f. Resultado de añadir a una
cantidad otra u otras homogéneas. 6. f. Operación matemática de sumar.
/// <>Suma<>:
www.elabueloeduca.com - (12/03/2017 - Domingo. 18:48);
La operación de sumar es la
primera de las operaciones fundamentales de la aritmética. Se representa con el
símbolo "+".
Consiste en dado un número añadir (adicionar) el valor de otros. Por
eso esta operación se llama también Adición.
Los números que vamos añadiendo se llaman sumandos y el resultado
obtenido se denomina suma o total. Continua en "elabueloeduca.com"
<>Sumar<>:
rae; Del latín medieval·summare·, y este del latón ·summa·'suma'.
1. tr. añadir (// agregar). Usado también como pronominal, A su falta
de tiempo se suma ahora una nueva obligación. 2. tr. Reunir en una sola
varias cantidades homogéneas. 3. tr. Dicho de varias cantidades: Componer una
total. 4. tr. desusado. Resumir, compendiar, abreviar una materia que estaba
extensa y difusa.
·Sumidero:
Rae; De ·sumir· y ·-dero·. 1. m. Conducto o canal por donde se sumen las aguas.
·Suministrar
Rae; Del latín ·subministrâre·.
1. tr. Proveer a alguien de algo que necesita.
·Sumir:
Rae; Del latín ·sumêre·.. 1. tr. Hundir o meter debajo de la tierra o del agua.
2. tr. sumergir (//abismar, hundir).
<>Superíndice<>:
rae; De ·super-· e ·índice·.
1. m. Letra o número que se coloca en la parte superior derecha de un
símbolo o de una palabra para distinguirlos de otros semejantes.
·Surco:
rae; De ·sulco·. 1. m. Hendidura que se hace en la tierra con el arado. 2. m.
Señal o hendidura prolongada que deja una cosa que pasa sobre otra. 3. m. Arruga en el
rostro o en otra parte del cuerpo. 4. m. Cada una de las estrías de un disco musical.
(A mi entender es un surco continuo decreciendo hacia el centro del disco).
a surco 1. loc. adj. Dicho de dos labores o hazas: Que están contiguas o
solo surco por medio.
<>·Surgir<>:
rae; Del latín ·surgêre·. 1. intr. Dicho del agua: Surtir, brotar hacia arriba. 2. intr. Aparecer o manifestarse, brotar. 3. intr. Dicho de una nave: Dar fondo.
<>Sustancia también substancia<>:
rae; Del latín ·substantia·.
1. f. Materia caracterizada por un conjunto específico y estable de propiedades.
Sustancia viscosa, orgánica. 2. f. Parte esencial o más importante de algo.
No traicionaba la sustancia del pacto firmado. 3. f. Conjunto de características
permanentes e invariables que constituyen la naturaleza de algo. La palabra
democracia está haciendo en las sociedades una evolución social hacia la convivencia
pacífica y en libertad. 4. f. Valor, importancia o utilidad de algo.
Un discurso con poca sustancia 5. f. Jugo extraído de un alimento. Se
aprieta la cebolla para que dé todo su sustancia. 6. f. Valor nutritivo de
un alimento. La leche materna supera en sustancia a la artificial. 7. f.
coloq. Juicio o sensatez. Una persona sin ninguna sustancia. 8. f. Fil.
Realidad que existe por sí misma y es soporte de sus cualidades o accidentes.
9. f. Chile. Golosina preparada con claras de huevo batidas, azúcar y gelatina,
de consistencia esponjosa y forma de bloque pequeño rectangular, cubierto con
azúcar de flor. 10. f. Nicaragua. Consomé. 11. f. Paraguay. Alimento elaborado
con leche, huevo y azúcar, que se da a personas convalecientes. 12 f. desusado.
Hacienda, caudal, bienes.
/// <>Sustancia simple<>:
WikipediA - (29/01/2017-Domingo.19:37);
Una sustancia simple , es aquella formada por moléculas de una sola clase de átomos. Por ejemplo, el dioxígeno, más conocido como oxigeno (O2), y el ozono (O3) son sustancias simples, porque sus moléculas están formadas sólo por átomos de oxígeno. Otro ejemplo lo constituye el diamante y el grafito. Que son sustancias simples por estar formadas por átomos de una única clase, los del elemento carbono, etc.
Lo contrario a una sustancia simple es una sustancia compuestas o compuesto. Los noventa y dos elementos químicos neutros se combinan entre sí formando casi tres millones de sustancias compuestas, denominadas compuestos químicos o, simplemente, compuestos.
·Sustrato también substrato:
Rae; Del latín ·substratus·, participio pasivo de ·substernêre· "extender bajo el suelo",
"poner como lecho".. 1. m. Estrato que subyace a otro. 2. m. Biol. Lugar que
sirve de asiento a una planta o un animal fijo. 3. m. Bioquím. Sustancia sobre
la que actúa una enzima. 4. m. Fil. sustancia (-- características permanentes de algo)
(-- realidad que existe por sí misma). 6. m. Fotogr. Baño aplicado al soporte para
permitir la adherencia entre la capa sensible a la luz y el vidrio o las materias
plásticas. 7. m. Geol. Terreno situado debajo del que se considera. El sustrato
del manto. 8. m. Ling. Lengua que ejerce una influencia en otra instalada
posteriormente en el mismo territorio. 8. m. Ling. Influencia que ejerce una
lengua en otra instalada posteriormente en el mismo territorio.