
T - t
·Tabique:
rae; Del árabe hispano ·tasbik·, y este del árabe clásico ·tasbïk· 'acción de
enrejar'. 1. m. Pared delgada que sirve para separar las piezas de la casa.
2. m. División plana y delgada que separa dos huecos. El tabique de las fosas
nasales.
·Tabla:
Rae; Del latín ·tabula·. 1. f. Pieza de madera plana, de poco grueso y cuyas
dos caras son paralelas entre sí. 11. f. Lista o catálogo de cosas puestas por orden
sucesivo o relacionadas entre sí. La tabla periódica de los elementos químicos.
12. f. Cuadro o catálogo de números de especie determinada, dispuestos en forma
adecuada para facilitar los cálculos. Tabla de multiplicar, de logaritmos, astronómica.
·Tabla periódica:
Rae; 1. f. Fís. y Quím. Ordenación de los elementos químicos según su número
atómico y dispuestos de tal modo que resultan agrupados los que poseen propiedades
químicas análogas.
/// Tabla periódica de los elementos:
WikipediA (10/04/2016); La tabla periódica de los elementos es una disposición de
los elementos químicos en forma de tablas, ordenados por su número atómico (número
de protones), por su configuración de electrones y sus propiedades químicas. Este
ordenamiento nuestra tendencias periódicas, como elementos con comportamiento similar
en la misma columna.
Las filas de la tabla se senominan períodos y las columnas
grupos. Algunos grupos tienen nombres. Así por ejemplo el grupo 17 es el de los
halógenos y el grupo 18 el de los gases nobles. La tabla también se divide en cuatro
bloques con algunas propiedades químicas similares. Debido a que las posiciones están
ordenadas, se puede utilizar la tabla para obtener relaciones entre las propiedades
de los elementos, o pronosticar propiedades de elementos nuevos todavía no descubiertos
o sintetizados. La tabla periódica proporciona un marco útil para analizar el
comportamiento químico y es ampliamente utilizada en química y otras ciencias.
Dimitri Mendeléyev publicó en 1869 la primera versión de tabla periódica que fue
ampliamente reconocida. La desarrolló para ilustrar tendencias periódicas en las
propiedades de los elementos entonces conocidos, al ordenar los elementos basándose
en sus propiedades químicas, si bien Julius Lothar Meyer, trabajando por separado,
llevó a cabo un ordenamiento a partir de las propiedades físicas de los átomos. Mendeléyev
también pronosticó algunas propiedades de elementos entonces desconocidos que anticipó
que ocuparian los lugares vacíos en su tabla. Posteriormente se demostró que la
mayoria de sus predicciones eran correctas cuando se descubrieron los elementos
en cuestión.
La tabla periódica de Mendeléyev ha sido desde entonces ampliada
y mejorada con el descubrimiento o síntesis de elementos nuevos y el desarrollo de
modelos teóricos nuevos para explicar el comportamiento químico. La estructura actual
fue diseñada por Alfred Werner a partir de la versión de Mendeléyev. Existen además
otros arreglos periódicos de acuerdo a diferentes propiedades y según el uso que
se le quiera dar (en didáctica, geología, etc.).
Se ha descubierto o sintetizado
todos los elementos de número atómico del 1 (hidrógeno) al 118 (ununoctium); la IUPAC,
Unión Internacional de Química Pura y Aplicada, confirmó los elementos 113, 115, 117 y
118 el 30 de diciembre de 2015. Los primeros 94 existen naturalmente, aunque algunos
solo se han encontrado en cantidades pequeñas y fueron sintetizados en laboratori antes
de ser encontrados en la naturaleza. Los elementos con números atómicos del 95 a 118 solo
han sido sintetizados en laboratios. Allí también se produjeron numerosos radioisótopos
sintéticos de elementos presentes en la naturaleza. Los elementos del 95 a 100
existieron en la naturaleza en tiempos pasados pero actualmente no. La investigación
para encontrar por síntesis nuevos elementos de números atómicos más altos continúa.
·Talo:
Rae; Del griego · ????? thálos· "retoño", "rama joven". 1. m. Bot. Cuerpo
de las talofitas, equivalente al conjunto de raíz, tallo y hojas de otras plantas.
·Talofita:
rae; Dicho de una planta: que tiene como cuerpo vegetativo un talo, que puede
estar constituido por una sola célula o por un conjunto de células dispuestas en
forma de filamento, de lámina, etc., ejemplo las algas o los hongos.
/// Talofítico:
Universidad de Sevilla - http://www.aloj.us.es/ - (28/11/2016-Lunes); En los niveles
de organización morfológica de los eucariotas: Son los individuos pluriceluares poiquilohídricos
con células +- diferenciadas por su función. Con Células reproductoras +- diferenciadas.
·Tamaño, ña:
rae; Del latín ·tam· 'tan' y ·magnus· 'grande'. 1. adj. Tan grande o tan pequeño.
Tamaña insensatez. 2. adj. Muy grande o muy pequeño. 3. m. Mayor o menor volumen
o dimensión de algo.
<>Tangente<>:
rae; Del antiguo participio activo de ·tangir·; lat. ·tangens·, -entis.
1. adj. Que toca. 2. adj. Geom. Dicho de dos o más líneas o superficies: Que
se tocan o tienen puntos comunes sin cortarse. 3. f. Geom. Recta que toca a una
vurva o a una superficie sin cortaleas. 4. f. Geom. Cociente entre el seno y el
coseno de un ángulo.
<>Tangente de un ángulo<>:
rae;
1. f. Mat. Tangente del arco que le sirve de medida.
/// <>Tanto por ciento <>:
www.profesorenlinea.cl - (23/01/2017-Lunes);
El porcentaje o tanto por ciento (%), es una de las aplicaciones más usadas de las proporciones o razones.
El porcentaje es un forma de comparar cantidades, es una unidad de referencia que relaciona una magnitud (una cifra o cantidad) con el todo que le corresponde (el todo es siempre el 100), considerando como unidad la centésima parte del todo.
Ejemplos:
1 centésimo = 1/100
5 centésimos = 5/100
50 centésimos = 50/100
Nota importante: No olvidar que las fracciones deben expresarse siempre lo más pequeñas posible, deben ser fracciones irreductibles.
¿Qué significa 50%?: Significa que de una cantidad que se ha dividido en cien partes se han tomado 50 de ellas, o sea, la mitad.
¿Qué significa 25%?: Significa que de un total de 100 partes se han tomado 25, o sea 1/4 (25/100 al simplificar por 5, se reduce a 1/4).
·Taxón, también Taxon,:
rae; Palabra creada sobre taxonomía 1. m. Biol. Cada una de las subdivisiones
de la clasificación biológica, desde especie, que se toma como unidad, hasta el
filo o tipo de organización.
/// <>·Taxón<>:
WikipediA - (06/01/2017-Viernes-18:17);
En biología, un taxón o taxon (del griego t????, transliterado como
táxis, «ordenamiento») es un grupo de organismo emparentados, que en una
clasificación dada han sido agrupados, asignándole al grupo un nombre en latín,
una descripción si es una especie, y un tipo. Cada descripción formal de un taxón
es asociada al nombre del autor o autores que la realizan, los cuales se hacen
figurar detrás del nombre. En latín el plural de taxón es taxa, y es como suele
usarse en inglés, pero en español el plural adecuado es «taxones» o «táxones».
La disciplina que define a los taxones se llama taxonomía.
La finalidad de clasificar los organismos en taxones formalmente definidos
en lugar de grupos informales, es la de proveer grupos cuya circunscripción
(esto es, de qué organismos están compuestos) sea estricta y cuya denominación
tenga valor universal, independientemente de la lengua utilizada para la
comunicación. Nótese que los taxones existen dentro de una clasificación dada,
sujeta a cambios y sobre la que pueden presentarse descrepancias; lo que obliga,
respecto a ciertas denominaciones problemáticas, a especificar en el sentido de
qué autor se está usando el nombre.
·Taxonomía:
rae; Del griego ·t???? táxis·'ordenación' y -nomía. 1. f. Ciencia que trata de
los principios, métodos y fines de la clasificación. Se aplica en particular, dentro
de la biología, para la ordenación jerarquizada y sistemática, con sus nombres, de
los grupos de animales y de vegetales. 2. f. Clasificación (// acción de clasificar).
<> · Taxonomía <>:
Para está página web o tejido informático: "software = compuintangible",
"hardware = compudáctilo".
WikipediA - 20/11/2017, 16:54, Lunes.
La taxonomía (del griego [τάξις - "táxis" -
'ordenamiento'] y [νόμος - "nómos" - 'norma' o 'regla'])
es, en su sentido más general, la ciencia de la clasificación. Habitualmente se
emplea el término para designar a la taxonomía biológica, la «teoría y práctica
de clasificar organismos». Como se la entiende en la actualidad, la clasificación biológica
tiene que ser congruente con las hipótesis de árbol filogenético disponibles, en
ella, los organismos se agrupan en taxones mutuamente excluyentes a su vez agrupados
en taxones de rango más alto también mutuamente excluyentes, de forma que cada
organismo pertenece a uno y sólo un taxón en cada rango o "categoría taxonómica".
En cada taxón, los organismos poseen caracteres (singular carácter) cuyos estados
del carácter los diferencian de los organismos fuera del taxón, y son los atributos
que en conjunto se utilizan para delimitar los taxones biológicos. La mayoría de
los especialistas ve a las especies, a las que ubican en taxones en la categoría
taxonómica del mismo nombre, como una realidad objetiva, y a los diferentes conceptos
de especie como diferentes aproximaciones para reconocer los linajes que evolucionan
independientemente, por lo que a pesar de las discrepancias entre autores en su concepto
taxonómico y por lo tanto en general en los límites taxonómicos de cada una, suele
ser considerada la categoría taxonómica más importante en la clasificación. Los
taxones supraespecíficos son los que agrupan especies y se jerarquizan en categorías
taxonómicas de rango cada vez más alto: género (grupo de especies), familia, etc.,
hasta el reino. En esta área también está debatido el concepto de taxón a utilizar.
Muchos especialistas afirman que sólo la "filosofía taxonómica" para agrupar los
organismos en taxones de la escuela cladista logra que la clasificación ofrezca el
servicio que se espera de ella al resto de las ramas de la biología, pero la
escuela evolucionista tiene razones para delimitar con otros conceptos los taxones,
también congruentes con las hipótesis de árbol filogenético, y las dos escuelas
se atribuyen la creación de sistemas de clasificación que cumplen en mayor medida
sus funciones de sistemas de almacenamiento y recuperación de datos, de
predictores del estado de los caracteres y las direcciones de la evolución que
no se han medido.
(22/11/2017, 15:54, Martes.)
La Taxonomia también se ocupa de debatir y actualizar los Códigos Internacionales
de Nomenclatura, de los que por razones históricas hay uno para cada disciplina
(Zoología, Botánica y Bacterias), cuya utilización para reglamentar el uso de
nombres "formales" está consensuada desde hace unos 100 años. Un Código define el
sistema taxonómico -los Códigos en uso definen el recién descripto, conocido como
el "sistema lineano", del que se deriva la definición de taxón más abajo descripta-;
decide cómo se forman los nombres y cuál es el "tipo" con el que se heredan de
taxón en taxón, y explicita sus Principios de Nomenclatura, cuya validez
está por encima de las reglas, con el objetivo o "principio básico" de proveer
la máxima estabilidad en la nomenclatura. Los Códigos en uso no evitan ls "diferencias
filosóficas" entre taxónomos en los conceptos taxonómicos a utilizar ni en la
categoría especie ni en las categorías supraespecíficas que son una fuente de
inestabilidad frecuente en los nombres de los taxones.
La Taxonomía Biológica es aquí tratada como una subdisciplina de la
Biología Sistemática, que además tiene como objetivo la reconstrucción de la
filogenia, o historia evolutiva, de la vida. Como aquí comprendida la Taxonomía
abarca la decisión de qué sistema taxonómico utilizar, la de qué conceptos taxonómicos
asociar a esos taxones, la delimitación de esos taxones dejando explícitos los
métodos que fueron utilizados para alcanzarla, la formalización de sus nombres
en la subdisciplina de la Nomenclatura, y también las herramientas para la
Determinación o identificación de especímenes. Como se calcula que sólo un
10% de las especies está descripto, esas descripciones tienen diferentes grados de
resolución, y las especies evolucionan (cambian con el tiempo en términos de sus
caracteres) y cambia su distribución, la determinación funciona recursivamente con
la descripción y catalogación de especies en el área llamada alfa-Taxonomía
o Taxonomia Descriptiva, o deslindándola de la definición minimalista de
Taxonomía que se maneja desde hace décadas y debido a que se intergradan en esa
categoría taxonómica, el área de la Sistemática Descriptiva o Ciencia
de Biodiversidad, la que se ocupa del descubrimiento, la identificación, la
descripción, la clasificación y la catalogación de la biodiversidad del planeta,
utilizando el organismo como unidad de trabajo y cuya prioridad es la descripción
de todas las especies dentro de un contexto evolutivo.
La historia de la taxonomía ha sido largamente recorrida por éstos y
otros interrogantes y a lo largo de ella fueron formándose el sistema de clasificación
y las reglas que nos han quedado como legado.
La crisis de biodiversidad que es foco de atención desde mediados de
1990, la cantidad de datos acumulados con los análisis de ADN, y la posibilidad
de intercambiar información a través de internet, han revitalizado a esta ciencia
en el ambiente científico desde las décadas de 1990-2000, y han generado un debate
acerca de la necesidad de hacer reformas sustanciales a los Códigos, algunas de
las cuales aún se están debatiendo y otras ya se han incorporado. Algunas de estas
propuestas son el BioCode, el PhyloCode, el agregado de atributos al tipo nomenclatural
como podrían ser los nuevos "marcadores de ADN" (ADN barcodes), y las
relacionadas con la informatización de los datos y la utilización formal de
Internet.
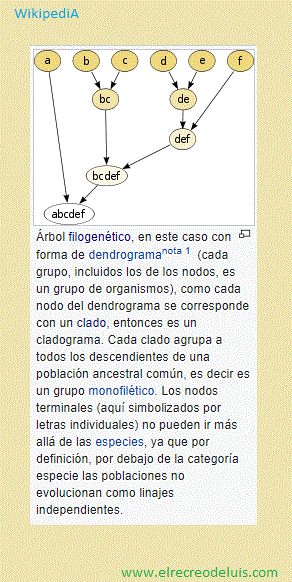

Definición de taxón
El taxón es un grupo de organismos con circunscripción, posición y rango.
De la estructura de la clasificación biológica como dada en la introducción, comúnmente
llamada "taxonomía linneana" o "sistema linneano" ,
(pero ver cita 9 =
"Taxonomía linneana" y "jerarquías linneanas" suelen ser los nombres utilizados en
referencia a esta taxonomía por autores como RR Sokal (1986), DL. Hull (1979), Judd
et al. (2007.548:" ...the use of Linnaean ranks..."), QD Wheeler (2008:6)"... a formal
Linanaean classification..." RK Brummitt "...Linnaean classification" etc. Pero ver:
Styessy (200026:232):
"-Comentario aparte, sería de ayuda si los defensores del PhyloCodedejaran
de referirse al sistema nomenclatural botánico moderno como el «Sistema Linneano»
(p.ej. Cantino 2000:87). Linneo delineó sus reglas de nomenclatura en Critica Botanica
(1737), que tiene poca resemblanza con nuestro moderno Código Internacional de
Nomenclatura Botánica (Greuter et al. 1994) desarrollado por mucha gente a través
de muchas versiones a lo largo de los últimos 130 años.)...
(24/11/2017, 15:30, Viernes.)
, se deriva la definición de taxón, que como explicitan los Códigos de Nomenclatura,
es un grupo de organismosque posee:
« circunscripción: el conjuto de caracteres obtenidos de los
organismos que lo componen que, junto con la regla de decisión propuesta al observalos,
diagnostican al taxón, establecen sus límites o lo delimitan, vuelven al taxón único,
diferencian sus organismos claramente de los organismos fuera del taxón único; la
circunscripción se define en función de poder determinar si un organismo nuevo pertenece
o no al taxón mediante la observación de sus caracteres según el protocolo dado,
y ella puede incluir idealmente o no una lista de caracteres diagnósticos (presentes
en ese taxón y sólo en ese taxón), el término aristotélico "definición" se sigue utilizando
en el Código de Zoología para referirse a la descripción exhaustiva que debería hacerse
de un taxón al nombrarlo, de la que se estraería la circunscripción;
« un rango o jerarquía (absoluta) llamado categoría taxonómica;
« y una posición taxonómica formada por los taxones de rango más
alto que lo contienen.
El taxón se redefine según el "sistema" taxonómico del que se deriva,
si bien la circunscripción es necesaria para ubicar organismos nuevos en su respectivo
taxón y por lo tanto, fundamental en todas las clasificaciones. Así, puede ser
modificada su definición para adaptarse a taxonomías que no utilizan rangos, o
pueden definirse los taxones de formas más restringidas, pero la filosofías
taxonómicas que han formado tres escuelas de la sistemáticas que dejaron un legado
a la materia, se diferencian en el concepto que debe utilizarse para aglomerar los
organismos en taxones, es decir su definición conceptual, su delimitación conceptual,
el concepto que relaciona los organismos del taxón más entre sí que con los demás
organismos, los diferentes "conceptos de taxón" o "conceptos taxonómicos".
La taxonomía según las diferentes escuelas
Según la escuela cladista, la que predomina hoy en día, la taxonomía es
la ciencia que debe decidir qué clados del árbol filogenético serán representados
por taxones y en qué categoría taxonómica ubicarlos.
Si bien en los libros de texto el clado en general se define como "una
secuencia de una población ancestral y todos sus descendientes", resulta ser una
definición poco rigurosa: el clado, como lo utilizan los taxónomos, agrupa sólo
poblaciones vivientes o al menos las que se utilizaron para tomar los datos, las que
en el método original de Hennig -el fundador, o la "voz más articulada" del cladismo-
se asocián como descendientes de un mismo linaje cuando se "encuentran" o determinan sus
apomorfías: los cambios de carácter en el linaje ancestral. Otras formas de análisis
filogenético como los métodos fenéticos (cuyo punto de partida es ciego en lo que
respecta a la evolución de los caracteres) fueron incorporados a la definición de clado,
de forma que pueden hipotetizar un "clado" (poblaciones agrupadas por poseer
caracteres compartidos desde el ancestro común más reciente) mediante métodos
de análisis no Hennigianos. El lector interesado puede recurrir a las siguientes
lecturas, listadas en Judd et al. (2007): Farris (1979), Donoghue y Cantino (1988),
Nelson y Platnick (1981), Humphries y Parenti (1986), Brooks y Mclennan (1991),
Forey et al. (1991), Cracraft y Donoghue (2004).
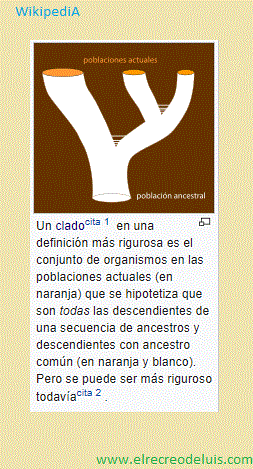
"Clado" proviene de cladogénesis. El conjunto de todos los descendientes
de un linaje se llama grupo monofilético (mono: único, phylum: linaje)
sensu Hennig, y los términos usualmente son utilizados como sinónimos. Los
taxones que representan grupos monofiléticos se llaman "taxones monofiléticos", y
las apormofías del clado que representan, que pueden haberse perdido o vuelto a
derivar en algunos miembros del grupo, pueden presentar variación fenotípica en
diferentes ambientes en los que se presenta el grupo que debe ser expresada, pueden
presentar los mismos genes pero otras variaciones debido a interacciones con otros
genes en diferentes individuos o grupos que también debe ser expresada, y pueden
se compartidas por otros clados que las hubieran adquirido independientemente.
Sólo después de haber obtenido una circunscripción (con apomorfías con toda su
variación fenotípica y genotípica, las plesiomorfías necesarias) y decidido una
categoría taxonómica, se puede considerar al grupo un taxón "lineano" y decidir si
nombrarlo según las reglas formales de nomenclatura.

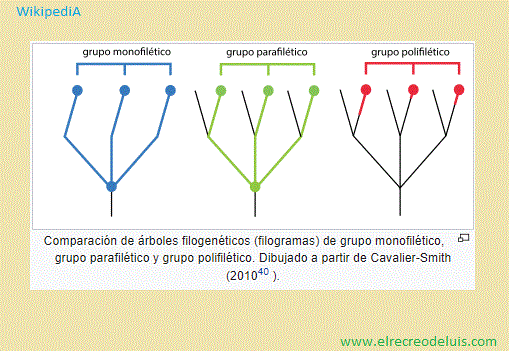
Hay otras escuelas de clasificación. Quizas la más importante, dentro
de las "minoritarias" es la que considera que los grupos parafiléticos también
deberían ser representados por taxones nombrados formalmente, si los grupos que
los conforman son lo suficientemente similares entre sí y los suficientemente,
disímiles del clado que queda afuera (escuela evolucionista). Los autores listados en
Judd et al. 2007 para el lector interesado: Simpson 1961, Ashlock 1979, Cronquist
1987, Mayr y Ashlock 1991, Stuessy 1983, Stevens 1986. Ejemplos de "taxones parafiléticos"
que se utilizan en las "clasificaciones evolucionistas" son los procariotas
(parafiléticos con respecto a los eucariotas) en el clásico sistema de clasificación
en reinos iniciado por Whittaker (1969 -Margulis 1971) y que hoy se sigue utilizando
(p.ej. Cavalier-Smith 1998), y las dicotiledóneas, parafiléticas con respecto a las
monocotiliedóneas (p.ej. Takhtajan 2009).
Otra escuela que fue encabezada por RR Sokal y PHA Sneath (Sokal y Sneath 1963,
Sheath y Sokal 1973, comentarios de los autores originales en Sokal 1986 y Sneath 1995)
recordaba que como no había forma de testear si eran verdaderas las "hipótesis previas"
sobre evolución, los razonamientos se hacían circulares: los "grupos preferidos"
determinan los "grupos preferidos". En los '60 cada autor de una clasificación proponía
un árbol completamente distinto y eran pocos los grupos que se mantenían entre
autores. Sokal y Sneath no sólo presentaban novedosos métodos de análisis "numéricos"
-cuantitativos- que sacaban provecho de las en ese momento novedosas computadoreas,
sino que también eran métodos "fenéticos": intetaban no asumir hipótesis de evolución
para armar el árbol, que debía buscarse siempre que existiera la posibilidad. En
las últimas décadas, en que "toda la sistemática es en alguna forma numérica"
debido a la cantidad de caracteres que se obtienen a su vez de cantidad de líneas
de evidencia diferentes (a las tradicionales se sumaron los análisis moleculares de
ADN, los caracteres bioquímicos y de ultraestructuras), se pudo dar vida útil a los
métodos de esta escuela fenética que al mismo tiempo perdió visibilidad, porque a
medida que se agregan hipótesis evolutivas para aumentar la congruencia entre líneas
de evidencia diferentes, los taxónomos van llegando a un consenso acerca de cuáles
serán las "verdaderas" de ellas con las que estimar la filogenia, con las que inferir
las relaciones cladísticas. Desde mediados de los '90 la sistemática vivió momentos
emocionantes: se empezaba a vislumbrar un único árbol predictivo y robusto, y
efectivamente, muchos grupos fueron abandonados por no estar basados en hipótesis
evolutivas correctas, incluidos algunos de larga data que causaron mucha sorpresa.
Los autores de esta tradicionalmente llamada "escuela" enfatizaban que sólo propusieron
métodos de análisis numérico, que fueron sólo una parte de importantes avances
numéricos en análisis filogenético, sistemática molecular, análisis morfométrico
e identificación númérica.
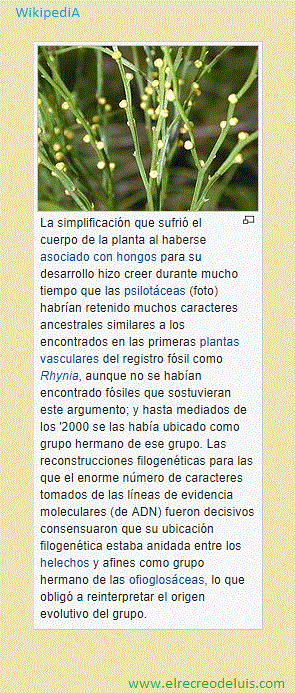
Hoy, las clasificaciones cladistas predominan por sobre las evolucionistas.
Las dos escuelas admiten que se necesitan nombres tanto para los grupos parafiléticos
como para los monofiléticos que los abarcan, la discusión entre cladistas y
evolucionistas radica en cuál es el sistema de nombres que debería formalizarse
en una clasificación. Los evolucionestas formalizan nombres de grupos parafiléticos,
y mencionan con nombres informales los grupos monofiléticos no clasificados
(por ejemplo Cavalier-Smith 1998 en adelante formaliza los reinos y nombra los
supergrupos de eucariotas sin formalizarlos), mientras que los cladistas formalizan
sólo grupos monofiléticos, llamando con nombres informales los grupos parafiléticos
(por ejemplo Al et al. 2005 formalizan los supergrupos de eucariotas y
proponen el nombre informal procariotas o bacterias para ese grupo parafilético).
Las dos funciones de los sistemas de clasificación
Más allá de los resultados de los análisis filogenéticos, todas las
escuelas coinciden en que la formalización de taxones en sistemas de clasificación
tiene dos funciones, en miras de ser un servicio para todo un abanico de usuaarios
finales, que no siempre pueden hacerse coincidir.
« Deben permitir hacer identificaciones, o visto de otra manera,
deben funcionar como un sistema de almacenamiento y recuperación de datos;
para lo que cada taxón debe estar claramente delimitado: un espécimen encontrado
del que se pudieran obtener los estados de sus caracteres, debería poder ubicarse
inequívocamente en un taxón conocido. Los científicos en todas las disciplinas deben
utilizar los taxones como unidad de trabajo, y publicar los resultados de sus
trabajos en relación al taxón estudiado. Por lo tanto los nombres científicos de los
organismos, los taxones formales deben representar el consenso de la comunidad
científica, sólo de esta forma funcionan como llave de acceso a un cuerpo de
información que refiere siempre al mismo grupo de organismos, aunque esté disperso
en libros y revistas escritos en muchas lenguas por diferentes autores, o preceda
de diferentes campos de la biología o la divulgación: como idealmente hacen
referencia los taxónomos, "El nombre de un organismo es la llave hacia su literatura.
« Deben permitir hacer predicciones, o pronósticos (que a diferencia de las
predicciones no están ligados a un método sino a un autor, en este caso el autor
de la clasificación y quizás también el autor final) y dar explicaciones. Por
ejemplo, cuando se encuentra un compuesto de interés médico en una planta, es
común que hay interés en investigar si ese compuesto se encuentra también en
otras especies relacionadas con ella; una taxonomía bien armada debería permitirle
al investigador presuponer a priori los límites de su investigación para
los que sea más probable confirmar sus hipótesis y ahorrar esfuerzos de muestreo.
La decisión de si armar la clasificación para predecir o para identificar
recorrió toda la historia de la taxonomía; las dos funciones coincidirían en una
clasificación que fuera al mismo tiempo "natural" como "artificial", es decir cuyos
conceptos taxonómicos sean "naturales" y cuyos taxones sean "artificiales" -como
siempre lo son, en las definiciones aquí manejadas-, con claves de identificación
que la reensamblen en grupos con parecidos superficiales y por lo tanto muy poco
predictivos, pero con reglas de decisión explícitas con las que se llegue con facilidad
al taxón terminal. La dificultad radica en que si se hace el ejercicio de buscar un
solo carácter, que sea diagnóstico de un taxon, (si el organismo posee un estado
del carácter, pertenece al taxón, si posee otro estado, no pertenece al taxón,
invariablemente, es decir una clasificación monotética), se notará que esta búsqueda
puede fallar al menos potencialmente en todos los grupos "naturales", debido a que
todos los caracteres son inestables en algún grado y pueden evolucionar, y otros
pueden presentarse como polimorfismos y el mismo estado coalescer de forma de
encontrarse en parte de taxones diferentes, y esta situación también se encuentra
si se toma un conjunto de caracteres. La forma alternativa de circunscripción es
proponer los estados de forma probabilística, esto es, un organismo es miembro de
un taxón si posee un número muy alto de estados de caracteres similares a la
descripción del taxón, especialmente de los poco lábiles. El mismo algoritmo politético
dará siempre el mismo resultado, basado en las probabilidades, pero a veces puede no ser el
eperado. Aquí se encuentra la difrencia fundamental entre lo que los autores llaman
clasificaciones "naturales" y "artificiales": en la clasificación natural, la determinación
es una probabilidad de estar acertado, dicho de otra manera un porcentaje de veces
se espera que la determinación llegue al taxón correcto pero no al concepto taxonómico
esperado. Esto es porque el grupo (el concepto taxonómico) existe independientemente de si
el taxónomo lo circunscribió correctamente, e independientemente de la clave de
identificación utilizada. En las clasificaciones artificiales, con tanto taxones
como conceptos taxonómicos artificiales, la determinación, aunque provenga de un
pesaje probalilístico de los caracteres, siempre es la correcta, aunque el grupo
no resulte ser lo que el taxónmo tenía en mente. El trabajo de taxónomo se manifiesta
en su habilidad para corregir los grupos artificiales creados por el hombre
(los taxones) de forma de acercarlos cada vez más a los grupos naturales (los
conceptos "naturales") con cada agregado de información nueva.
(25/11/2017, 11:37, Sábado.)
Si un grupo "natural" es un grupo "predictivo" y esto, en taxones
superiores a especie, lo logran el número y peso de los caracteres, medidos y no
medidos, compartidos por los miembros del grupo que evolucionan juntos, entonces la
"naturalidad" de los grupos será una cuestión de grado, y habrá grupos más o menos
similares, o más o menos "naturales". Su componente más importante es el histórico
o filogenético, que además ofrece impresas en sus similitudes fenéticas las
restricciones construccionales que restringen las direcciones de la evolución del
grupo de una fomra que no puede ser explicada por factores ecológicos. Tanto
cladistas como evolucionistas se adjudican las clasificaciones con mayor valor predictivo,
o "más naturales" (p.ej. T Cavalier-Smith, E. Mayr, NI Platnick, Hörande y Stuessy).
Construyendo una clasificación
Hoy en día los taxónomos, aun dentro de la escuela cladista (p. ej. Judd
et al. 2007:33), tienen claro que no es de utilidad que cada clado o concepto
de taxón por arriba de especie se formalice, es decir se corresponda con un
"taxón nombrado" acorde a las reglas consensuadas en los Códigos internacionales
de Nomenclatura. Las decisiones a la hora de sintetizar el árbol, inevitables,
pueden parecer un poco arbitrarias por falta de métodos confiables para lograr los
dos objetivos recién mencionados, pero se pueden generalizar algunos criterios. (:32-35):
1. El criterio principal es la fuerza de la evidencia que respalda al
grupo: para convetir un grupo en taxón formal debe haber un análisis filogenético
que lo respalde de forma robusta, sobre la base de más de una línea de evidencia
de forma que todas ellas muestren congruencia. Según Judd et al. esto se
evidencia en una cantidad importante de apomorfías que definan su ancestro.
2. Debe haber una cantidad de caracteres diagnósticos que permitan
diferenciarlo claramente del resto de los taxones. Según algunos sistemáticos cada
taxón debería poder ser diagnosticado morfológicamente en lo posible (p.ej. «18»,«104»...)
3. Deberían poder inferirse aspectos de su biología únicos a ese taxón,
para ello debería tener caracteres morfológicos interesantes ("obvios") que permitan
que el taxón se comporte como una unidad predictiva (Judd el al. 2007:33)
4. Algunos autores afirman que los taxones deberían tener, en lo posible,
entre tres y siete subtaxones, un número que puede manejar con facilidad la memoria
humana. En las palabras de Davis y Heywood (1963. 83 citado en Judd et al. 2007):
"Debemos ser capaces de ubicar a los taxones en taxones de categoría más alta de
forma que podamos encontrarlos de nuevo". Los sistemas de clasificación permiten la
creación de categorías intermedias hasta lograr el número de taxones deseados en cada
una.
5. Por estabilidad de la nomenclatura, los taxones que ya han sido
nombrados en el pasado deben continuar con el mismo nombre en lo posible, a menos
que haya fuertes justificaciones para abandonarlos.
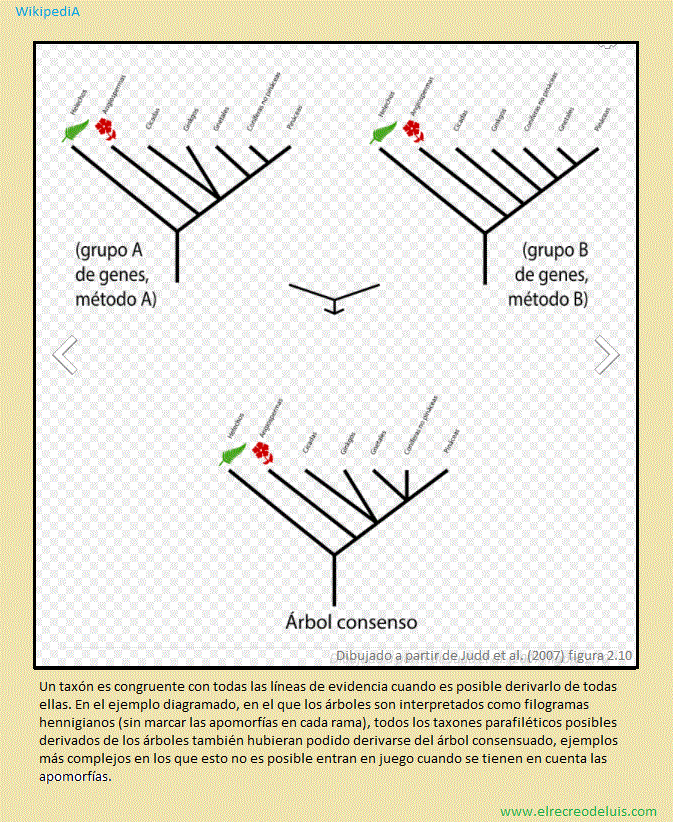
Hay dos criterios para decidir la categoría taxonómica. Uno, el tradicional,
es el que existe desde los inicios de la clasificación y es anterior a la teoría
de la evolución; éste decide la categoría taxonómica según el grado de variación
del plan corporal entre taxones. Desde la incorporación de la teoría de la evolución
al pensamiento científico, se interpreta a la categoría taxonómica como la que
deriva de un grupo de apomorfías que aportan cierto grado de variación en el plan
corporal del taxón en ralación a los demás taxones en la misma categoría. Entonces
los reinos y los filos (las categorías más altas) derivan de transformaciones
evolutivas importantes que definieron de nuevo el plan corporal, a las que siguió
un batallón de novedades evolutivas forzadas a adaptarse al nuevo cambio, mientras
que las categorías más bajas, como género y especies, poseen variaciones en caracteres
más triviales, y por lo tanto más inestables. Esta construcción de categorías,
basada en variación de caracteres más a menos estables, es la que llegó hasta nuestros
días y probó ser útil para los usuarios finales de una clasificación (ej. Cavalier-Smith
2010). Otro criterio de clasificación en categorías fue el iniciado por Hennig
(1950, 1966), quien proponía que la edad geológica absoluta de un grupo fuera el
criterio para decidir su categoría taxonómica, y si bien hay científicos trabajando
para lograr esta meta en algunos grupos según otros parece ser un concepto abandonado.
Las categorías resultaron ser útiles para los objetivos de la clasificación (ej.
«108»,«109»,«110»), pero algunos autores las desestiman por considerarlas arbitrarias
y hasta los hay que proponen eliminar el sistema de clasificación con categorías
(por ejemplo en de Queiroz 2006). Por razones históricas el sistema definido por
los Códigos en uso utilizan las categorías linneanas de clasificación: reino, filo
o división, clase, orden, familia, género y especie.
La clasificación en la categoría especie
Un "concepto de taxón" particular es el de especie. La mayoría de los
especialistas ve las especies, a las que ubican en taxones en el rango o categoría
taxonómica del mismo nombre, como un "taxón natural", una realidad objetiva, haciendo
uso los diferentes conceptos de especie de las diferentes aproximaciones o las
diferentes propiedades emergentes en el nivel de organización del linaje que
evoluciona independientemente, esto es que, al menos potencialmente, adquiere su
propio set de cambios evolutivos. A diferencia de la situación en los rangos más
altos, el hecho de que este rango es objetivo y que existe al menos entre o en
forma de dos cotas superior e inferior es poco cuestionado en la práctica, le es
intuitivo al lego, y es el concepto que hace que cobren sentido objetivos de otras
ramas como "conservar todas las especies": un linaje que se pierde no puede ser
regenerado sin cambios evolutivos en linajes emparentados. Pero como no se sabe
cómo van a evolucionar los linajes y por lo tanto cuáles de las delimitaciones
posibles constituyen una "unidad evolutiva" en proceso de especiación, o cuál
considerar la "unidad evolutiva especie" cuando diferentes caracteres evolucionan
de a grupos de tamaños diferentes, la delimitación a veces se vuelve enteramente
una cuestión de consenso.
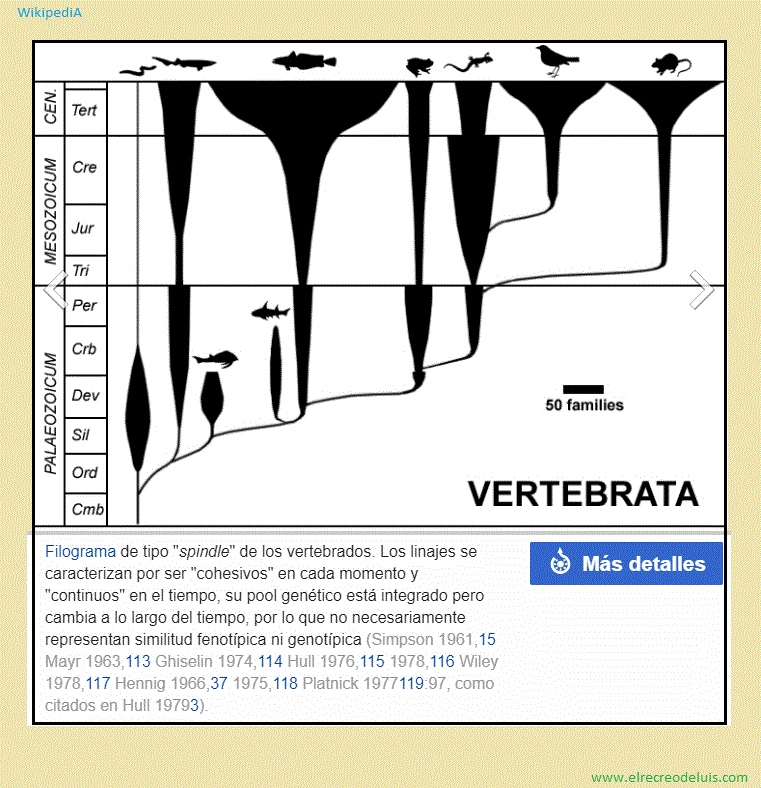
Un linaje, una especie, consta de una o un grupo de poblaciones, que
también son difíciles de definir, si bien el concepto espacial, sus límites
geográficos, metodológicamente siempre están presentes. Normalmente se definen
como un grupo de individuos de una misma especie que ocupan una región geográfica
más o menos bien definida y con los individuos interactuando entre sí. Estas
"interacciones" no sólo refieren al intercambio de genes que puede haberlo o no
según su tipo de reproducción sino también al hecho de que compiten por los mismos
recursos, comparten las mismas condiciones, son afectados por los mismos gremios
de predadores, patógenos y parásitos y de competidores interespecíficos por algún
recurso, poseen las mismas interacciones de simbiosis y comensalismo con otras
especies, y se reproducen y dispersan por los mismos mecanismos, por lo que la
población sufre además de los avatares del azar una selección de genes mediante la
supervivencia y reproducción sexual o asexual de aquellos individuos que los porten
y cuya descendencia eventualmente ocupa el nicho que ocupaban los demás. (El nicho
en ecología se define en función de la supervivencia del linaje: las características
ecológicas del género o rangos superiores no alcanzarían para explicar cómo sobreviven
sus linajes; algunos taxónomos pueden utilizar una definición más amplia de nicho
para referirse a los espacioes demensionales que acotan el nicho de las especies
involucradas en taxones superiores, concepto que otros prefieren llamrar "zonas
adaptativas".) Estas interacciones entre individuos que mantienen integrado el
linaje en un nicho diferenciado son definidas tradicionalmente como "cohesión", un
concepto con más factores que la posibilidad de hibridar en simpatría -el "concepto de
especie biológica" de Mayr-, pero que no siempre delimita unidades discretas: la
cohesivas y quizás a diferente grado en cada aspecto a evolucionar, aun sin corredores
los individuos de las poblaciones no se aparean al azar y se aparean o interactúan con más
probabilidad con los individuos más cercanos a sí mismos, por lo que priorizar
factores diferentes de la cohesión puede dar resultados diferentes. Incluso si se
llega a un acuerdo en estos factores y grados, si los linajes constan de un
continnum espacial que correlacione con un continnum de cohesión,
todavía pueden ser divididos en unidades de forma arbitraria. Esto llevaría a
clasificaciones incompatibles, cuando sus delimitaciones se solapan. En la búsqueda
de la categoría se puede reconocer una cota inferior
(una población que no sobrevive sin su metapoblación, es decir que no es independiente,
no es una especie) y una superior (por ejemplo, la descripción de un género no
debería alcanzar para explicar cómo sobrevive cada uno de sus linajes, cómo se
mantiene su "continuidad" en el tiempo). Esa búsqueda al menos hoy en día parece
ser infructuosa en algunos taxones, especialmente microbios de reproducción
asexual, cuya cohesión parece ser un continuum en el que todas las
categorías parecen ser arbitrarias.
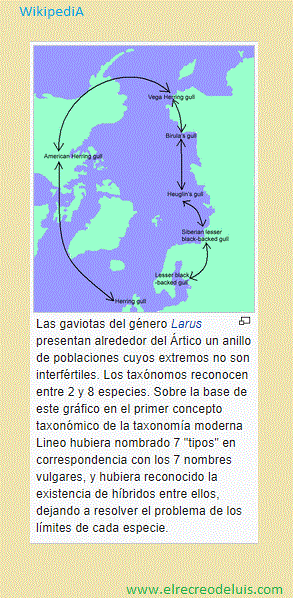
De los conceptos de especie, que son "casi tantos como especialistas
dispuestos a discutirlos", dos tuvieron importantes implicancias en la estabilidad
de la nomenclatura y en las ramas de la ciencia que los utilizan. Uno es el
"concepto de especie biológica" o BSC (biological species concept), que
delimita a la especie como el grupo de poblaciones que puede hibridar entre sí y
dar descendencia fértil, que no subdivide las comunidades reproductivas a menos
que se mantengan aisladas en simpatría, y es el concepto más utilizado, o al menos
el más invocado, desde su articulación por E Mayr en los 1940-1960 hasta los 1980-1990;
el otro que ganó popularidad en las últimas décadas es el "concepto de especie
filogenética" o PSC (phylogenetic species concept), que divide las comunidades
reproductivas en un grano más fino según si conservan caracteres en un análisis
de filogenia, y en muchos taxones, incluso en algunos que parecían muy estables
como Primates, se superpuso su clasificación a la BSC.
Una de las intenciones del BSC como formulado por E. Mayr era en parte
minimizar la confusión causada por la multitud de ecotipos dentro de las especies
biológicas. Esto es porque la práctica habitual es que se describan las especies
de una pequeña región dada, que en esa región están bien aisladas reproductivamente
entre ellas, pero en los intentos de integrar la información entre regiones es
común encontrar un continuum de tipos intermediarios. Sin embargo, el BSC,
fuera de los taxones de vertebrados de tamaño grande a los que Mayr estaba acostumbrado,
no resultó delimitar taxones con tanta precisión como parecía, entre las especies
biológicas puede haber diferentes grados en la capacidad de hibridar y quizás en
un 10% de las especies biológicas de animales, un 25% de las especies biológicas
de plantas y un porcentaje más alto de microbios se encuentren híbridos y no
necesariamente con especies hermanas; por lo que se ha denominado "divisores"
(splitters) y "agrupadores" (limpers) (misma terminología que
en taxones superiores:238-241), mientras que en los continuum sigue
existiendo el problema de los solapamientos. Habiendo decidido utilizar
ese concepto de especie, todavía se suman a la incertidumbre las inconsistencias
históricas entre la delimitación práctica -normalmente fenética- y la mucho más
costosa delimitación conceptual de las especies. El requerimiento clave para
solventar estas diferencias es que los taxónomos sean explícitos acerca de qué
concepto taxonómico y qué método para alcanzar su delimitación están siendo
utilizados, de forma de poder superponerlas y utilizarlas sincronizadamente cuando
eventualmente se encuentra más de una clasificación describiendo la misma región.
Nomenclatura biológica
El último paso en una clasificación consiste en buscar qué nombre
científico le corresponde al taxón o averiguar si se necesita publicar uno
nuevo, y eso se hace según las reglas escritas en los Códigos Internacionales de
Nomenclatura. Por razones históricas hay uno para cada disciplina: de Zoología,
de Botánica, y de Bacterias, y se actualizaban frecuentemente como resultado de
los Congresos Internacionales que reúnen a los científicos para tal efecto. Por
ejemplo, a julio del 2015 el último Código de Botánica es el de Melborne (revisión
2011, publicado en 2012), el último de Zoología es la cuarta edición (revisión
1999, publicado en el 2000), y el último de Bacterias es la revisión de 1990
(publicado en 1992). Las ediciones pueden no ser la última versión, ya que se
han publicado posteriormente enmiendas. Los Códigos poseen algunas diferencias entre
sí, pero en su esencia funcionan de forma similar. Con el objetivo, y el
"principio básico", de la "estabilidad en la nomenclatura" los autores de los
Códigos enumeran sus principios, de los que el "Principio de Prioridad"
consiste en que cada taxón con su circunscripción, posición y rango debe poseer
el primer nombre de los establecidos para él como único "nombre aceptado" ("nombre
correcto" en Botánica, "nombre válido" en Zoología) para la comunicación.
El principio de prioridad es sencillo de expresar, lo dificultuoso es
decidir cuándo dos autores se están refiriendo al mismo taxón y cuándo a uno diferente,
o dicho con más rigor, los dificultuoso es decidir, cuando un nuevo autor cambia
las hipótesis taxonómicas de forma de recircunscribir los grupos o cambiarlos de
posición o rango, cuál de los nuevos taxones deberá utilizar un nombre ya utilizado
en el pasado. Los Códigos deciden que el nuevo taxón que "hereda" el nombre es el
que contiene el mismo tipo dentro de él. Los Códigos buscan que cada descripción,
cada concepto taxonómico o cada conjunto de subtaxones que ha ya sido nombrado
continúe con el mismo nombre en lo posible. De esta forma los Códigos definieron
lo que en nomenclatura se llama el "taxón nominal", la asociación permanente entre un
"nombre" y un "tipo", este último establecido, en estos Códigos, en una categoría
taxonómica primaria dada. Esta asociación perdura a través de los cambios de
circunscripción, necesaria para establecer y reutilizar el taxón nominal pero
variable entre autores; a través de los cambios de posición taxonómica, que en la
categoría especie lleva a una modificación en el nombre; y a través de los cambios
de categoría si se cambia a una categoría cercana que mantiene el tipo (del mismo
"grupo de categorías"), que normalmente lleva también alguna modificación en el
nombre.

El "tipo", el contenido del taxón que se fija permanentemente al nombre,
es algo diferente si estamos hablando de un nombre en la categoría especie o inferior,
o de un nombre de una categoría superior a especie. En los Códigos en uso, en la
categoría especie o inferior a especie (las categorías del grupo especie), "el
espécimen tipo nomenclatura" es un espécimen específico de la especie idealmente
completo, en buenas condiciones, y depositado en una colección cuya manutención y
crecimiento sean dirigidos por un taxónomo especializado (un curador), quien
debería permitir al público un acceso a los tipos aunque sea restringido. Cuando
es posible, los tipos se depositan en instituciones dedicadas a la ciencia como
museos de historia natural y jardines botánicos. Cuando el tipo utilizado en la
publicación original (el "holotipo") se ha perdido o vuelto irreconocible, no hay
otro espécimen de la colección original que pueda reemplazarlo (un "lectotipo"),
y hay necesidad de un tipo para reutilizar un nombre, el nombre no se pierde sino que
se puede seleccionar de los especímenes identificados con el mismo nombre y localidad
que el original, un "neotipo". En los casos particulares de bacterias, protozoos,
algas y hongos cuyas colecciones constan de cepas vivas, el tipo no es un único
organismo muerto sino que es una cepa cultivada viva, en general congelada, y
disponible en al menos una o dos colecciones diferentes según el Código.
Con respecto a los taxones superiores a especie, el nombre del género -y
de las demás categorías del grupo género- posee como "tipo" al taxón nominal de
aquella especie contenida en él que fue publicada primero; y el nombre de una
familia -y de las demás categorías del grupo familia- posee como "tipo" al taxón
nominal del género contenido en ella que fue publicado primero. Nótese que en
estos casos el "taxón nombrado" es un grupo de organismos cuyo tipo
es un género o una especie contenidos en él, aunque, como está nombrado según las
reglas formales de nomenclatura, el tipo contiene a su vez un espécimen tipo o un
taxón que eventualmente lo anida. Con los cambios de circunscripción, ese espécimen
es el único que se mantiene necesariamente dentro de los límites de cada taxón de
cualquier categoría.
Si dentro de la extensión (la composición de organismo) del taxón no
hay ningún ejemplar especificado como "tipo", el autor de la circunscripción puede
decidir nombrarla formalmente, es decir, crear un taxón nominal que
pasará al terreno de los "nombres científicos" por el resto de la historia de la
taxonomía. El tipo será un ejemplar identificado dentro de la extensión del
taxón, debidamente depositado en una colección, y el nombre será un nombre en latín
o latinizado construido según ciertas reglas explicitadas en los Códigos (algunas
de las cuales se verán más abajo). Todos los datos del nuevo taxón (el nombre, la
posición, la categoría taxonómica, la descripción o la diagnosis escrita en forma
de caracteres expresados en palabras, los datos de la ubicación del tipo) son
"efectivamente publicados" si lo son donde sean accesibles para otros científicos
y expertos locales, como una revista científica, o un libro. No se consideran
efectivamente publicados los nombres publicados en medios efímeros de publicación
como un catálogo de semillas, un diario, o por correo electrónico. Todos estos
datos efectivamente publicados convierten al taxón nominal en un nombre establecido
("válidamente publicado" en Botánica "disponible" en Zoología), y es cuando decimos
que un nombre tiene el estatus de "nombre científico" o de "nombre formal".


Cuando se encuentra más de un "tipo" dentro del taxón, los demás nombres
pasan a ser sinónimos taxonómicos del primero, el "nombre aceptado" ("nombre correcto"
en Botánica, "nombre válido" en Zoología) del taxón. Pero si al descubrirse este
hecho un nombre más reciente ya está muy extendido, puede proponerse a la
Comisión respectiva como excepción al principio de prioridad y en aras de la
estabilidad, agregar el nombre a la lista de nomina conservanda, nombres que
se consideran el nombre aceptado por razones prácticas.
Los nombres también pueden ser establecidos pero ilegítimos. Esto
es porque las reglas enunciadas para establecerlos no han impedido su
formación, pero se descubre que no cumplen con los principios, los que se
intentan cumplir mediante las reglas y que son enunciados en la primera parte de
cada Código. Por ejemplo los principios más importantes para mantener la estabilidad
en la nomenclatura son que debe haber un solo nombre para cada taxón, y el principio
de prioridad. Las dos razones principales por las que un nombre puede ser ilegítimo
son que al momento de la publicación no se ha evitado que (a) fuera redundante,
es decir que el autor establece un nombre científico para un taxón que en la
composición de organismos posee un ejemplar que ya forma parte de un taxón nominal
y es así de una forma "evidente", y (b) que fuera homónimo -no sinónimo- de
otro anterior, es decir que un "nombre" que ya formaba parte de un taxón nominal fuera
asociado a un "tipo" nuevo. Mientras no haya un registro central de nombres
incluido en las reglas estas situaciones no son evitables mediante ellas, y a pesar
de que se lo intenta no son todavía fáciles de detectar, se publican miles de
nombres nuevos por año y sólo se ha logrado establecer un registro central
mandatario de nombres y publicaciones en bacterias, que existe desde 1980. No se
han terminado de informatizar los datos de la literatura publicada en papel para
indexarlos en bases de datos digitales, y botánicos y zoólogos no consensúan la
creacción de "índices de nombres adoptados" ("no suprimidos", dejando los
nombres no indexados suprimidos por defecto) en los taxones que les tocan por
temor a que unos pocos especialistas dejen publicaciones fuera de la indexación
y acaparen la nomenclatura de un grupo.
Cuando la aplicación estricta de un Código resulta en desestabilización
de la nomenclatura, o no cumple los Principios, como se argumenta cuando se realizan
propuestas de conservación de nombres o de supresión de nombres pobremente establecidos,
la situación se preseta ante su Comisión respectiva, que tiene la responsabilidad
de tomar una decisión al respcecto. La composición y funciones de cada Comisión
están contepladas en su respectivo Código, y tanto las propuestas como las decisiones
tomadas en relación a ellas son publicadas en su respectiva revista especializada.
Las decisiones tomadas por la Comisión Internaciona de Nomenclatura Zoológica son
publicadas en su revista. The Bulletin of Zoological Nomenclature, las
tomadas por los Comités auspiciados por la Asociación Internacional para la
Taxonomía de Plantas (IAPT) son publicadas en su revista, Taxon, y las
tomadas por el Comité Internacional de Sistemática de Procariotas Y Bacteriología
son publicadas en su revista, el international Journal of Systematic and Evolutionary
Microbiology (IJSEM).
Finalmente, la nomenclatura botánica, la nomenclatura zoológica y la
nomenclatura bacteriológica son independientes la una de la otra, (con pocos taxones
exceptuados). Como consecuencia, el mismo nombre puede ser utilizado para una
planta, para un animal o para una bacteria, sin que sean formalmente nombres
homónimos, y aunque no es lo aconsejable hoy en día animales y plantas pueden
estar compartiendo el 13% de sus nombres en la categoría de género.
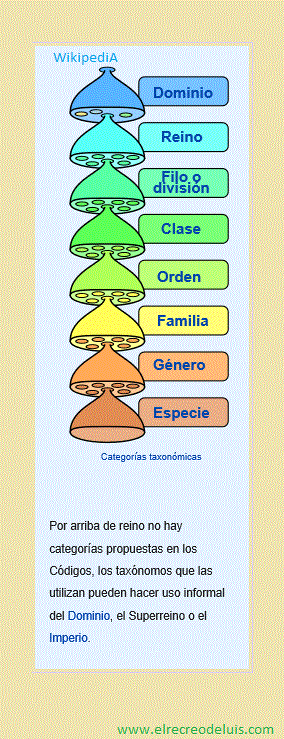
Categoría taxonómica
En los Códigos de Nomenclatura, son siete las categorías primarias o
principales: Reino, Filo o División, Clase, Orden, Familia, Género, Especie.
Históricamente, el Código de Zoología utilizó la categoría Filo (Phylum en latín
y en inglés), y el Código de Botánica y el de Bacterias escindido de él, la de
División (Divisio en latín, Division en inglés), hoy en día en
una dirección hacia la armonización se los trata como sinónimos.
Una regla mnemotécnica para recordarlas es la siguiente:
El rey es un filósofo de mucha
clase que ordena para su
familia, géneros de buena especie.
- Reino, Filum, Clase, Orden, Familia, Género, Especie.
La reina es una diva de mucha
clase que ordena para su
familia, géneros de buena especie.
- Reino, División, Clase, Orden, Familia, Género, Especie.
El establecimiento de un nombre en una categoría primaria genera un
"tipo" en lo que los Códigos llaman el "grupo de categorías" que abarca todas las
categorías intermedias por arriba y por debajo de la categoría primaria, que
poseerán, justamente, el mismo "tipo". Estos taxones serán establecidos cuando
se los publique por primera vez, con sus respectiva circunscripción.
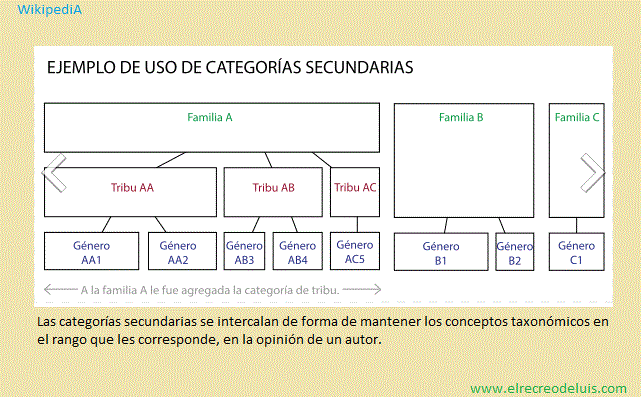
Luego de establecer las categorías primarias pueden intercalarse algunas
categorías secundarias, pero a diferencia de las primarias éstas son optativas, y
no es necesario intercalarlas a lo largo de toda la categoría en una clasificación,
pero una vez que se decidió agregar una categoría secundaria a un taxón primario,
sí es necesario agregarla a lo largo de todo ese taxón. Son tribu entre familia y
género), variedad y forma debajo de especie, y las que utilizan prefijos: superfamilia,
subfamilia. La categoría de subtribu sólo puede establecerse después de la de tribu,
y la de subsección después de la sección. Las categorías secundarias propuestas por
cada Código se presentarán en la sección siguiente.
(26/11/2017, 11:37, Domingo.)
Nombre científico
En los Códigos de Nomenclatura, cada especie queda designada por un
binomio (una expresión de dos palabras) en latín, donde la primera palabra, el
"nombre de género", es compartida por las especies del mismo género; y la segunda,
el "adjetivo específico" o "epíteto específico", hace alusión a alguna característica
o propiedad distintiva de esa especie en particular, como pueden ser el color
(albus, 'blanco'; cardinalis, 'rojo cardenal'; viridis,
'verde'; luteus, 'amarillo'; purpureus, 'púrpura'; etc.), el origen
(africanus, 'africano'; americanus, 'americano'; alpinus,
'alpino'; arabicus, 'arábigo'; ibericus, 'ibérico'; etc.), al hábitat
(arenarius, 'que crece en la arena'; campestris, 'de los campos';
fluviatilis, 'de los ríos'; etc.), un homenaje a una personalidad de la
ciencia o de la política o atender a cualquier otro criterio. No es necesario que
el nombre esté en latín, sólo es necesario que esté latinizado. Los nombres de
género siempre van con la primera letra en mayúsculas, los epítetos específicos
siempre van en minúsculas, y los nombres de género y de especie van siempre en
itálicas (o subrayados, si se escribe a mano). Al escribir el nombre de especie,
el epíteto específico nunca es utilizado solo, y es mandatario que esté precedido
por el nombre del género, de forma que el nombre de la especie sea el binomio completo.
La abreviación a la primera letra del nombre del género precediendo el epíteto específico
completo también es aceptable una vez que el nombre ya apareció en su forma completa
en la misma página o en un artículo pequeño. Así por ejemplo, la lombriz de tierra
fue llamada Lumbricus terrestris por Linneo, y si el nombre ya apareció antes en el
artículo y no hay ambigüedad, puede volver a refefrirse a ella como L. terrestris.
Con respecto a los taxones ubicados en la categoría de género y superior, los
nombres son uninomiales (constan de una sola palabra) y siempre se escriben con
la primera letra en mayúsculas (aunque solamente en la categoría de género van en
itálicas). El epíteto específico debe concordar gramaticalmente con el género, y
si en su género original no concordaba se modifica para que lo haga: Leucadendron
argenteum fue originalmete descripto en otro género como Protea argentea.
Como los Códigos de Nomenclatura prohíben que dentro de cada Código haya dos taxones
con el mismo nombre, es decir dos taxones nominales con el mismo nombre, no puede
haber dos géneros con el mismo nombre (ni dos taxones por arriba de género con el
mismo nombre), pero como ocurre que el epíteto específico de las especies sólo se
usa después del nombre del género, puede haber dos especies diferentes pertenecientes
a géneros diferentes que compartan el mismo epíteto específico. Una vez
fijado, un nombre no es sustituido por otro sin un motivo nomenclatural. Por ejemplo
el roble de los alrededores de Madrid fue bautizado como Quercus pyrenaica
erróneamente, puesto que no se encuentra en los Pirineos, pero tal circunstancia no
justifica un cambio de nombre.
La recomendación es que los nombres sean eufónicos, fáciles de memorizar
y descriptivos, y a que los lectores aprendan a obtener ese atractivo con el
objetivo de que sean utilizados por una parte significativa de los usuarios
finales, dedicó Linneo muchas de sus secciones en Philosophia Botanica
1751); pero no hay reglas que impidan a los investigadores utilizar su propia
inventiva para crear los nombres científicos más insólitos: el nombre de la araña
Pachgnatha zappa, porque tiene una mancha en el abdomen igual al bigote
del artista Frank Zappa; algunas moscas chupadoreas de sangre del género Maruina:
Maruina amada, M. amadora, M. cholita, M. muchacha, M. querida, M. chamaca,
M. chamaguita, M. chica, M. dama, M. nina, M. tica y M. vidamia, todos
adjetivos cariñosos; el dinosaurio que fue llamado Bambiraptor debido a
Bambi, el de la película de Disney; el molusco bivalvol Abra cadabra
(aunque después se lo cambio de género); y quizás el caso más sobresaliente, el
género de arañas brasileñas Losdolobus, llamado así porque los investigadores
que lo describieron, queriendo homenajear a dos argentinos que habían colaborado
en parte significativa en la publicación de la que no iban a ser los autores, les
pidieron a éstos que inventasen un nombre para el nuevo género, que quedó como
Losdolobus por "los dolobus", término del lunfardo argentino intraducible
en una enciclopedia. Los Códigos en una de sus recomendaciones hacen un llamado a
la responsabilidad, "Los autores deberían ejercitar un razonable cuidado y consideración
en la formación de nombres nuevos para asegurarse de que son elegidos con sus
subsecuentes usuarios en mente y que, mientras sea posible, sean apropiados, compactos,
eufónicos, fáciles de memorizar y no cusen ofensa.
En las categorías superiores a género los nombres científicos están
compuestos por la raíz del tipo (un género del taxón, en los rangos más
bajos el más antiguo) seguida de un sufijo que denota la categoría en la que el
taxón fue ubicado, como se indica en la siguiente tabla:
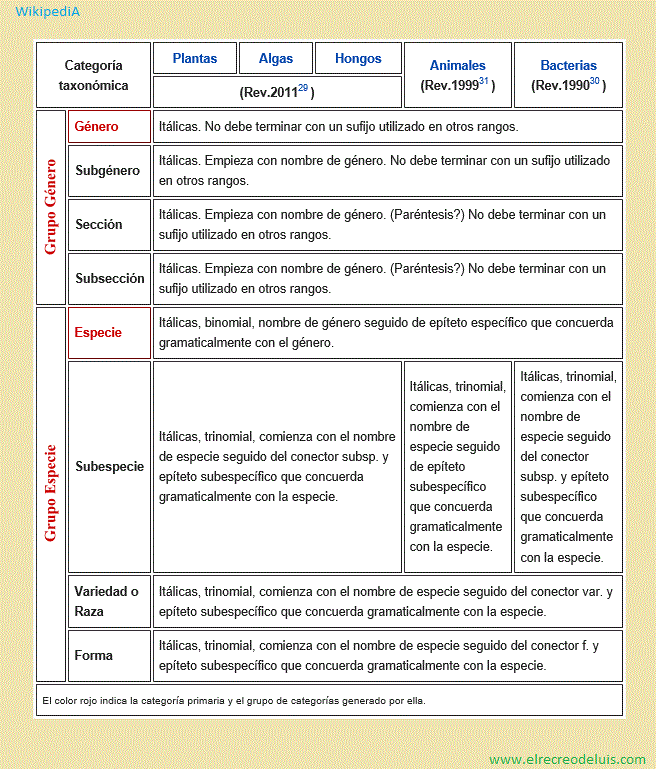
Además, en todos los taxones, la primera vez que se nombra un taxón en
una publicación científica, el nombre científico puede verse seguido del apellido
del autor de su primera descripción, que es el que estableció el taxón nominal
(normalmente llamado la "autoridad"), mejor aún si se sigue del año en que el
taxón fue establecido. El nombre del autor puede estar abreviado, por ejemplo a
Linneo se lo abrevia con una «L.». Las veces posteriores que se nombra al taxón
en la publicación se evita el autor y el año de publicación. A veces se encuentran
variaciones en la forma en que el autor y año son escritos (por ejemplo hay dos
autores conectados por palabras en latín como in o et o ex,
o el autor está entre paréntesis), y cada una de estas variaciones tiene su
significado en Nomenclatura. Por ejemplo, en el caso de los animales, si en la
actualidad una especie en particular está ubicada en un género diferente al que le
fue asignado por su autoridad original, el nombre del autor y el año se ponen
entre paréntesis indicando así que originalmente se la describió con el mismo
taxón nominal, pero otro nombre. Así, el león en las clasificaciones actuales es
«Panthera leo (Linnaeus, 1758)», y se pone autor y año entre paréntesis porque
Linneo lo describió originalmente como «Felis leo» (Linneo seguía el nombre del
taxón del autor de su concepto taxonómico, que no siempre coincidía con el primer
autor del nombre). El autor y año de publicación no son reglas sino recomendaciones
de los Códigos de Nomenclatura, pero son de uso corriente y se encuentra en la
literatura taxonómica que son altamente recomendados por muchos autores. Cuando se
hace referencia a la circunscripción de un concepto taxonómico utilizado en la
actualidad que difiere de la circunscripción de la publicación del taxón nominal,
se puede encontrar el conector sensu, p.ej. «Arenaria sensu McNeill (1962)».
El autor y año del taxón nominal original se mantienen útiles para evitar errores
con nombres homónimos. Esperablemente, en el futuro esta notación pueda reemplazarse
de forma de desambiguar todos los conceptos taxonómicos asociados al mismo nombre.
En informática de la biodiversidad, se propone utilizar un Identificador Global
de Conceptos Taxonómicos (de nombres según cada autor), que al resolverse
guíe a la cita del concepto utilizado con todas sus referencias y quizás un
enlace a la publicación y a los usos previos del nombre hasta el momento en que
fue establecido.
¿Por qué cambian los nombres de los taxones?
La descripción formal y la tipificación de los taxones son procedimientos
en los que es frecuente observar errores e inconsistencias, y esto ocurría aun en
mayor medida antes de que los Códigos Internacinales reglamentaran estos trabajos
muy pormenorizadamente. Los nombres de los taxones pueden cambiar cuando son
redescubiertas descripciones más antiguas del mismo taxón, y se aplica el principio
de prioridad, o cuando se observa que la descripción original en realidad refería
a un taxón diferente. No suelen darse estos casos en los grupos ya bien estudiados,
y en los Códigos hay además mecanismos para la supresión de nombres pobremente
definidos en los grupos más complicados. Una vez asentada la taxonomía fundamental
de un grupo, es poco probable que los cambios de nombres se den por esta clase
de motivos.
Los nombres cambian cuando el descubrimiento de nueva información obliga
a modificar las circunscripciones, pero también -y sin necesariamente nueva información
que lo justifique-, los nombres cambian por diferencias de criterio entre los
especialistas.
Por ejemplo, es muy común que a distintos grupos de organismos recién
descubiertos se les dé nombres de especie (y de género icluso) nuevos y que años
después un taxónomo que los estudie llegue a la conclusión de que todos pueden
hibridar entre sí y dar descendencia fértil, y unifique todos los taxones en una
única especie (en este caso todos los nombres cambian al nombre de la primera especie
descrita dentro del grupo), dejando, en todo caso, los antiguos epítetos de especies
como subespecies. Con posterioridad puede llegar otro taxónomo que considere que
esos grupos de organismos deberían dividirse en dos especies distintas, que en la
naturaleza conviven sin hibridarse (aunque pudieran), debido a que cada una de las
especies que él propone forma una unidad monofilética en un análisis de filogenia.
Entonces a los que se encuadran en la "nueva" especie, se les cambia el nombre de
especie por el nombre de la especie más antigua descrita dentro de la "nueva"
especie. Otro taxónomo podría argumentar más tarde que todos los grupos originalmente
descritos son especies válidas del mismo género, debido a que cada uno de ellos
puede ser identificado por su propio grupo de caracteres morfológicos, y que dentro
de cada uno de ellos hay un patrón de ascendencia y descendientes. En este caso,
todos los nombres de género se unificarían en uno solo, por lo que todos los
géneros pasarían a nombrarse como el género más antiguo descrito dentro del grupo.
Esta inestabilidad de los nombres se habría producido casi únicamente por cambios
de "filosofía taxonómica": el "concepto taxonómico" que cada autor considera que se
corresponde con la categoría especie. Estas diferencias conceptuales entre taxónomos
son extremedamente comunes hoy en día, lo cual lleva a una gran inestabilidad potencial
en los nombres de los taxones (Vane-Wright 2003 citado en Mallet y Wilmott 2003).
Determinación o identificación de especímenes
La disciplina de la Determinación, o identificación de especímenes,
es el proceso de tomar los atributos de un espécimen hasta encontrar el taxón en
el que se ubica ("determinarlo") o reconocer que es nuevo para la ciencia y necesita
una descripción formal y un nombre. Como sólo el 10% de las especies del planeta está
descripto, y esas descripciones son hipótesis con diferentes grados de resolución,
la Determinación funciona recursivamente con la descripción y delimitación de especies
en el área llamada alfa-Taxonomía o Taxonomía Descriptiva, o, modernamente
y deslindando las diferencias en las definiciones de Taxonomía -desde hace décadas
con una definición minimalista- y Sistemática, y debido a que se integraban en esa
categoría taxonómica, el área de la "Sistemática Descriptiva" o "Ciencia
de la Biodiversidad". Esta área se ocupa además de la investigación expedicionaria
y exploratoria en ambientes naturales para avanzar en el descubrimiento, la
identificación, la descripción, la clasificación y la catalogación de la biodiversidad
del planeta, utilizando como unidad de trabajo el organismo y cuya prioridad es la descripción
de las especies dentro de un contexto evolutivo, aproximándose a ello desde la
investigación primaria sobre un grupo taxonómico dado, un gremio ecológico, o
una región geográfica, se ocupa de realizar nuevas colectas (el "trabajo de campo")
y depositarias en colecciones, del trabajo de toma de datos sobre las colecciones
existentes, de la realización de clasificaciones revisionarias, de desarrollar
hipótesis taxonómicas (determinación de homologías, caracteres, apomorfías y plesiomorfías,
especies, clados), de testear las hipótesis taxonómicas de especie, y la creación
y uso de las herramientas informáticas necesarias para todo ello y para publicar las
nuevas claves de identificación y las nuevas descripciones, con las que en conjunto
se llega a los taxones redefinidos, que junto con el desarrollo teórico de las
hipótesis forman parte de la literatura taxonómica primaria, cuyos productos más
acabados son las llamadas floras, faunas y monografías taxonómicas. Durante una
determinación, todos los puntos tomados de la literatura deben ser comprobados con
los especímenes propios para tomarlos por verdaderos. Especialmente en áreas donde
la biodiversidad presenta una tarea desafiante, como en los trópicos -debido al
número de especies y también el vasto número de especies que no ha sido aún
publicado, o exhaustivamente fotografiado- el rol del especialista, profesional o
experto local, en la determinación es crítico.


Además de la finalidad en sí misma de generar una taxonomía sólida y
estable, hay varias otras razones por las que se puede necesitar una determinación,
y todas si están correctamente documentadas y los ejemplares depositados en
colecciones con acceso al público general y a la comunidad científica, aportan
datos a la alfa-Taxonomía. Los taxónomos necesitan identificar los organismos
sobre los que extraen nuevas líneas de evidencia con las que ajustar los análisis
de filogenia en taxones de categorías más altas, y a pesar de que se elige para sus
muestreos a los espcímenes que se consideran bien determinados tampoco ha faltado
algún error en la reconstrucción filogenética por un error de arrastre de determinación.
En Biogeografía se utilizan los datos de especímenes identificados por localidad u
"ocurrencias", lo que se llama "datos georrefenciados", en los análisis, cuyas
conclusiones a su vez pueden debilitar o robustecer hipótesis taxonómicas. Muchos
trabajos de investigación realizados por biólogos ecólogos implican un conteo
de alguna unidad taxonómica de biodiversidad en cierta área, como en las investigaciones
relacionadas con la conservación, o en las relacionadas con las invasiones biológicas
y su impacto en el ecosistema, o en los cambios en patrones de sucesión en relación a
algún otro tipo de disturbio. Una correcta identificación se necesita cuando se
sospecha que una enfermedad o muerte en humanos, animales o plantas es causada por
un ser vivo, como cuando se sospecha que el ganado está consumiendo una planta
tóxica que le produce enfermedad o muerte, o cuando un animal salvaje pica o muerde
a un humano, o un microbio patógeno lo infecta, la correcta identificación en estos
casos abre la literatura que indica las formas de control y tratamiento conocidas,
y estas ocurrencias agregadas pueden analizarse en la búsqueda de correlaciones y
patrones predictivos. Los fabricantes de pesticidas y herbicidas y los servicios
relacionados con la salud pública pueden requerir un monitoreo de insectos o plantas
dañinos en cada zona con el objetivo de mantener regulada su abundancia y para
probar efectividad de tratamiento cuando se decide apricarlos. En el campo de la
biología forense la identificación de especímenes esclareció asuntos legales.

Si se han realizado identificaciones para un trabajo que se está por
publicar, es buena práctica citar cuál fue el taxónomo consultado o las publicaciones
taxonómicas que fueron utilizadas pora identificar las especies, y agregar una
sección de Especímenes examinados donde se cite la ubicación de los
especímenes que fueron depositados o visitados en la colección que los preserva
como referencia (los boucher, «en tafalles 'recibo,vale'») de los datos
sobre los que se realizó la hipótesis publicada. La sección "especímenes examinados"
debería incluir el nombre científico actual [o el aceptado sensu qué autor],
el número de identificación del espécimen individual dentro de la colección
(el accesion, «en tafalles 'adhesión, acceso, entrada»), el nombre de la colección
o su acrónimo, la localidad donde fue colectado, y si se las obtuvieron, los
accessión de las secuencias de ADN publicadas por ejemplo en GenBank.
Si fue publicado el dataset «en tafalles 'conjunto de datos'» de
ocurrencias por ejemplo en GBIF, puede citarse la ubicación del mismo en el
repositorio público en el que fue compartido o las ocurrencias dentro del mismo
utilizadas. Las colecciones de especímenes poseen una infraestructura específica
para cada taxón, y cada autor puede tener una colección privada -incluso unicata-
(unicate collection), «en tafalles, 'colección unica'», con un solo
espécimen o lote, quizás holotipo o isotipo-, pero si es posible se alojan en
instituciones públicas como museos de historia natural o jardines botánicos, que
son los sitios más estables y las hacen perdurar por centurias, el taxónomo
especializado que dirige su manutención y crecimiento -el curador- idealmente
debería permitir al público un acceso a los especímenes aunque sea restringido.
Las colecciones suelen tener catálogos electrónicos con la información de los
especímenes que poseen, que son cada vez más fáciles de acceder y, a pesar de que
en sus inicios carecían de estandarización en sus formatos, se prevé que en el
futuro muchas de esas bases de datos puedan ser accedidas por un mismo portal,
esperablemente mediante los estándares TDWG. Uno de esos portales, creado en el
2001 por acuerdos internacionales entre agencias gubernamentales después de
la Convention on Biological Diversity (CBD), «en tafalles 'Convención sobre
diversidad biológica», es GBIF, que aloja tanto datasets «en tafalles
'conjunto de datos'» compartidos desde los catálogos de museos y herbarios como
datasets «en tafalles 'conjunto de datos'» que se publican asociados
a publicaciones científicas. En las colecciones, no es raro que los especímenes se
reclasifiquen si están los suficientemente bien documentados, se calcula que un
porcentaje que puede llegar a ser de hasta un 20% o un 25% de los especímenes puede
ser reclasificado durante el rechequeo «en tafalles 'recomprobar, reverificar,
recontrolar» (ver -179-); algunos de ellos podrán ser
designados como el "tipo" de un nombre de especie nueva o de una ya nombrada cuyo
tipo original se ha perdido. Son cada vez más comunes los sofwares «para mi página,
'compuintangible'» que formatean la
sección de especímenes examinados a partir de la planilla de datos, y también puede
formatearse en forma de data paper, «en tafalles 'papel de datos'»
un set «en tafalles 'conjunto, juego, serie, grupo, equipo.», de ocurrencias
para publicación formal.
Colecta
Sólo si las circunstancias lo permiten, porque pueden ser difíciles de
colectar o estar en peligro de extición, los especímenes a identificar pueden
colectarse con cuidado de no dañar ninguno de sus caracteres. Los especímenes no
tienen ningún valor si se pierde el dato de la localidad de la que se extrajeron
y el momento en que fueron encontrados, los datos de espécimen/localidad-momento
se llaman ocurrencias, sean éstos colectados, fotografiados o sólo ingresados
como dato (lo que puede permitirse según la confiabilidad del colector). Al colectar
el espécimen o al fotografiarlo se realiza una ficha de colección con datos
de localidad, fecha y colector; y otros que se perderán al llegar al lugar de
identificación como hábitad, si estaba asociado a alguna otra especie, su abundancia
en esa localidad, si era de noche o llovía, el método por el que fue colectado, y
cualquier información sobre el estado del espécimen que el colector piense que pueda
no ser evidente una vez llegue a su lugar de identificación: fotografiar o anotar
claramente el color y textura de los órganos -que podría cambiar debido a la preservación-,
anotar la fragancia, etcétera (18:553). Si se colecta más de un espécimen de especies
diferentes, debe hacerse una ficha de colección para cada ejemplar. Luego de la
identificación debe agregarse la misma a la ficha y también quién es el identificador,
que no necesariamente es la misma persona que hizo la colección, o cuáles fueron
los métodos utilizados para la identificación (claves o colecciones con las que
se lo comparó), para dejar lo más explícita que se pueda la crcunscripción utilizada
para ese mombre aceptado. Después de una campaña se posee típicamente para cada
espécimen, un id «en tafalles 'carné de identidad'», una ficha de colección
y un set «en tafalles 'conjunto,
juego, serie, grupo, equipo.» de fotos asociados a ese id «en tafalles
'carné de identidad'», una ubicación en la colección (una entrada o
accession, no todos los especímenes fueron colectados y poseen número de
accession, «en tafalles 'adhesión, acceso, entrada»), y hay entradas
que albergan un lote de varios organismos similares colectados en el mismo momento,
por lo que corresponden con varios id «en tafalles 'carné de identidad'»,
de especímen), y esperablemente también posee asociada una lista de los estados
de los caracteres que posee, necesarios para llegar a la identificación y que
fueron convenientemente dejados por escrito en alguna planilla estandarizada.
Cada id «en tafalles 'carné de identidad'» de espécimen representa una
fila en una colección de "ocurrencias". Si se ingresan las fichas en un formato
electrónico, debe preveerse que cumpla con los estándares provistos por el
Biodiversity Information Standards «en tafalles 'Estándares de información
sobre la biodiversidad'» (anteriormente TDWG) para facilitar
su acceso (ver el estándar para ocurrencias Darwin Core).
Identificación (Determinación)
El primer paso en una identificación es una observación a simple vista,
que implica una apreciación inconsciente de muchos caracteres del espécimen al mismo
tiempo, en la que se asocia el espécimen), los métodos más formales de identicación
entran en juego cuando el interesado necesita que la identificación continúe
hasta taxones más específicos, o cuando necesita confirmar una posible apreciación
(18:6-7,186). Los métodos formales tienen en cuenta pocos caracteres, expresados
además de forma "tipológica", resultando ser una simplificación de la apreciación
gestalt = """[un todo organizado que se percibe como algo más que la suma
de sus partes «forma» :
'en castizo -primera impresión secundada por la información- en castizo zarzuelesco
sería -estampación primigenia asistida por la averiguación-]"""
del taxónomo que los realiza; pero siempre que se dispone de ellos, es importante
seguirlos. Comparar directamente con las colecciones o las fotos puede inducir
errores, ya que en las fotos pueden no mostrarse o no ser evidentes para el lector
no entrenado los caracteres que diferencian el taxón de los demás.
Los taxónomos concuerdan en que la mejor forma (20:498) y la más rápida
(18:6-7) de identificar un espécimen con métodos formales es averiguar cuál es el
especialista, taxónomo o experto local, al que le son familiares las especies
de ese morfotipo o taxón superior en esa región y que tenga comprobada su existencia
en su propia colección y esté al tanto de las últimas actualizaciones publicadas
sobre el taxón. La determinación que él haga será muchas veces más segura que la
que el usuario final pueda hacer con la literatura y los instrumentos a los que
tenga acceso (20:498).
Los instrumentos para la identificación varían según el taxón
superior tratado. Para plantas y artrópodos, los instrumentos son lupa de disección,
y elementos para manipular la muestra en la lupa (dos agujas de disección y una pinza
de punta fina). en el caso de las plantas un bisturí para seccionar órganos o una
hoja de afeitar, puede ser útil un calibre (18:557) para hacer las mediciones, y
quizás saber hacer preparados para observar al microscopio (18:557). Si además
se tiene acceso a las fotografías del espécimen vivo o recién colectado la tarea
de identificación se ve facilitada. Raramente dos especies no pueden discernirse
morfológicamente y se debe recurrir al envío de muestras para la extracción y
análisis de ADN.


Las herramientas literarias para la comparación formal de esos
datos son las claves de identificación que hacen uso de los estados de los
caracteres del espécimen para descartar posibilidades hasta llegar al nombre
del taxón; de ellas el método mas difundido hoy en día es la clave dicotómica (18:557).
En la clave dicotómica se elige siempre uno de dos caminos, dos descripciones
mutuamente excluyentes de los estados de algunos caracteres. La elección envía al
usuario al siguiente par de descripciones, hasta que, si el usuario no cometió un
error, se llega al nombre del taxón (18:558,20;496-497). La tarea se ve muy facilitada
si la clave tiene fotografías del aspecto de los estados de los caracteres.

La clave se llama politómica si se dan más de dos opciones en
cada bifurcación, es una clave politómica en dos pasos la utilizada por Linneo en
Species Plantarum (1753). En el primer paso pedía contar el número de
estambres, y en el segundo el número de estigmas.
En la clave multi-acceso o policlave, se puede utilizar cualquier conjunto
de caracteres en cualquier orden, de forma que el usuario selecciona los caracteres
utilizados para la identificación. Estas claves normalmente contienen mucha más
cantidad de caracteres que las claves dicotómicas, y son muy ventajosas cuando se posee
un espécimen incompleto o cuando se tiene información incompleta sobre el espécimen.
Se han escrito programas de computadoras de claves multi-acceso, en que de forma
interactiva se ingresan los caracteres y el programa devuelve los posibles taxones.
Programas de identificación asistidos por computadora son por ejemplo el antiguo
sistema DELTA ("Descriptive Language for Taxonomy")(Askevold y O'Brien 1994, Dallwitz
1983, 2000, Dallwitz et al. 2002, Pankhurst 1978, 1991, Watson y Dallwitz 1991, 1993
citados en Judd e al 2007) (hoy superado por el TDWG estándar SDD), en la web se
pueden encontrar otros como el sistema LuciD (Norton 2012), y es seguro que se
volverán más importantes en el futuro. Los programas sirven tanto para utilizar
claves ya existentes como para construirlas a partir de una matriz de taxones
x caracteres, y están adaptados a que se agreguen fotos, videos y sonido. Hoy
en día, la mayor desventaja de las claves multi-acceso es su disponibilidad
(18:558,20:496-497), aunque ya hay que identifican las plantas de cualquier región
del mundo hasta familia (como en colby, discoverlife), hormigas (antweb), saltamontes
(Common Western Grasshoppers), mariposas y polillas (Butterflies and Moths), y
muchas todavía están siendo creadas (Farr 2006). Hay esfuerzos crecientes para
computarizar las claves de identificación, incluidas las multi-acceso.
Si bien las claves informáticas son valiosas, la mayor parte de las
claves de identificación todavía se encuentran, junto con las respectivas descripciones
e ilustraciones de cada taxón, en enormes volúmenes en papel publicado en series
llamadas floras y faunas, que reúnen la información sobre la presencia
de las plantas o animales pertenecientes a un área particular, a una pequeña región,
o a todo un continente (18:560). La primera flora de la taxonomía moderna,
Species Platarum (1753) de Linneo, reunía la información global hasta el
rango de especie y variedad en los conceptos taxonómicos del autor, totalizando unas
8000 especies; en la actualidad la flora global editada por Kubizki fue iniciada
en 1990, llega hasta el rango de género, y todavía no está terminda, los volúmenes
se siguen publicando esporádicamente 25 años después. Las floras y faunas son esfuerzos
conjuntos en los que un taxónomo editor le da a cada taxónomo o pequeño grupo de
ellos a describir exhaustivamente el taxón para el que es especialista, en general
hasta la categoría de especie en una región dada, convenientemente revisado, chequeado
con las colecciones existentes y la bibliografía previa convenientemente revionsada;
en general estos proyectos tardan décadas en completarse y pueden quedar imcompletos
si los subsidios en general gubernamentales dejan de renovarse. Un problema que deben
enfrentar los usuarios de estas claves es su antigüedad, por la que muchas veces
están desactualizadas. Los nombres recién descriptos para la ciencia pueden no encontrase
en ellas, como las nuevas revisiones de géneros y familias con especies exóticas que
se hayan instalado hace poco en la región. Otro problema es que dos floras o faunas
que afecten la misma área (como una flora o fauna local y una regional) pueden
diferir en sus conceptos taxonómicos o en sus circinscripciones y por lo tanto dar
resultados diferentes. Las claves computarizadas poseen muchos de los mismos problemas
que las claves en papel, aunque es esperable que en el futuro ofrezcan mejores
facilidades para ser actualizadas y para la búsqueda rápida de sinónimos. Las
actualizaciones se encuentran en las monografías (monographs), un
estudio extenso con claves, descripciones y fotos de un taxón en particular, en
general en el rango de familia o género, y en las revisiones taxonómicas
(taxonomic revisions), similares a las monografías pero menos exhaustivas.
Las dos últimas se publican en revistas científicas (18:560), y pueden ser accedidas,
como las floras y faunas, en jardines botánicos, museos y universidades con
bibliotecas y hemerotecas con acceso al público. Las guías de campo (field guides),
que todavía son más faciles de acceder y poseen muchas fotografías e ilustraciones
de las especies de un taxón en una región particular, se han vuelto muy populares
y son útiles como una primera aproximación a conocer la flora o la fauna de una
región, siempre que se utilicen con cautela y teniendo en cuenta que sólo describen
las especies más vistosas y no siempre pueden hacer uso de las herramientas que
diferencian un grupo de especies similares.
Al utilizar una clave, siempre hay que leer cuidadosamente las dos
opciones, tener en cuenta todos los caracteres mencionados, no estimar las
mediciones sino realizarlas, buscar en el glosario todos los términos que no se
conozcan, y tener en cuenta que los seres vivos poseen cierta variabilidad, así
que si existe la posibilidad de ver un carácter varias veces (por ejemplo en varias
hojas de una planta) se ganará en precisión (18:558). Muchas veces no es evidente
que una descripción se corresponde con el espécimen, porque no necesariamente todos
los caracteres de una descripción se corresponderán con todos los caracteres del
espécimen. Al utilizar una clave es muy probable que en algún momento se llegue
a un par de descripciones donde el camino a seguir sea confuso. Cuando es así,
hay que seguir los dos caminos, si alguno de los dos es muy incoherente (por
ejemplo si pide la observación de varios caracteres que no existen en el espécimen)
se descarta. Si no se pueden descartar caminos, se llegará a más de un posible
taxón (171,18:558,20:496-497). Aunque se obtenga un solo resultado, es importante
chequearlo. Una forma de hacerlo es leer la descripción y ver las ilustraciones
del taxón o los taxones a los que se llegó, especialmente evaluando la lista de sus
caracteres diagnósticos, que es una lista de todos los estados de caracteres
del taxón que lo sistiguen de los demás taxones. Si coinciden con los caracteres del
espécimen, entonces se ha realizado la identificación (18:558,20:495). Otras formas
de hacerlo son la comparación con alguna colección de especímenes, con colecciones
de fotos, o en especímenes difucultosos donde aún queden dudas, enviárselo a un
especialista mejor entrenado en ese taxón (20:497):
(27/11/2017, 16:55, Lunes.)
Historia de la taxonomía
Los orígenes de la taxonomía se remontan a los orígenes del lenguaje,
cuando las personas llamaban con los mismos nombres a los organismos más o menos
similares, sistema que persiste hoy en día en lo que llamamos los "nombres vulgares"
de los organismos. Todas las sociedades humanas poseen un sistema taxonómico que
nombra a las especies y las agrupa en categorías de orden superior. El ser humano
posee un concepto intuitivo de lo que constituye una especie, producto de agrupar
a los organismos según las propiedades emergentes observadas a ese nivel de
organización, y esto se ve reflejado en que todas las clasificaciones populares
de los organismos poseen remarcables similitudes entre ellas y también con las
utilizadas hoy en día por los biólogos profesionales.
La taxonomía biológica como la disciplina científica que conocemos hoy
en día nació en Europa. Alli, a finales de la Edad Media se habían formado las
primeras universidades, donde se discutían temas de índole filosófica y técnica.
Pronto los naturalistas se dieron cuenta de que, si bien los nombres vulgares son
útilies para el habla cotidiana, necesitaban un sistema más universal y riguroso
para nombrar a los organismos: cada especie debía ser nombrada, cada especie debía
poseer un único nombre, y cada especie debía ser descrita de forma inambigua.
Al finalizar la Edad Media, con la dominación militar y cultural que una Europa
revitalizada sostendría sobre el resto del mundo, el sistema allí consensuado se
extendería a todo el globo.
De la taxonomía linneana al origen de los Códigos.
En 1583, el italiano Andreas Caesalpinus habia delineado cuáles debían
ser las características de un sistema de clasificación: debía ser fácil de usar
y de memorizar, estable en el tiempo, conciso, y además predictivo (ver Greene 1983.
citado en Judd et al. 2007). Por lo tanto existía la conciencia de que un sistema
de clasificación no sólo debía reflejar la naturaleza (ser predictivo), sino también
servirle de utilidad a una comunidad de usuarios.
Por un tiempo los naturalistas hicieron intentos de ordenar la información
global sobre los organismos y reglamentar sus nombres. Pero sin duda alguna fue
en el siglo XVIII que la taxonomía recibión un empujon definitivo gracias al
nsturalista sueco Carlos Linneo, quien tenía la ambición de normalizar los nombres
de todos los animales, plantas y minerales agrupados según sus similitudes morfológicas.
Ya había publicado trabajos más modestos antes, pero no fue hasta 1753, ya largamente
pasados sus 40 años, que publicó una gigantesca Flora de dos volúmenes en la que
se encontraba ordenada toda la información dsiponible sobre las plantas (incluyendo
las sinomimias con los autores más relevantes de los no seguidos), y debido a su
éxito, terminó siendo el trabajo que definío el sistema de clasificación que se
utiliza hoy en día. El libro se llamaba Species Plantarum ("Las especies de
plantas", o "Los tipos de plantas"), y estaba escrito en latín, el idioma universal
de la época. En esta Flora Linneo agrupó sus "especies de plantas" en una categoría
de rango superior llamada género, llamados por él género "naturales", en los que
agrupaba las especies que pertenecían a ellos. Al describir una especie dentro de
cada género, sólo debía listar los caracteres de la misma que la diferenciaban de
las demás especies de su mismo género. Al enfatizar el uso de similitudes morfológicas
para agrupar las especies, Linneo estaba clasificando a los organismos en virtud
de sus similitudes genéticas, y sin haberlo conceptualizado de esa manera, por lo
tanto también evolutivas. En su libro, cada especie era descrita con una frase en
latín limitada a 12 palabras, en donde la primera siempre era el género al que
pertenicía la especie, y el resto los caracteres que las diferenciaban. A estas
pequeñas descripciones, o polinomios. él las llamó "nombre apropiado de cada
especie", pero hizo un importante agregado que había sido inventado en su momento
por Caspar Bahhin (1560-1624): la nomenclatura binomial o binomios, como
el "nombre corto" de las especies. En los márgenes del libro, seguido del nombre
"apropiado" polinomial de cada especie, escribió una sola palabras. Esta palabra,
combinada con la primera palabra del polinomio (todavía hoy llamada género), formaba
un nombre más fácil de recordar y corto para cada especie. Por ejemplo la hierba
de gato fue nombrada "apropiadamente" con el polinomio: Nepeta floribus interrupte
spicatus pedunculatis (en español, "Nepeta con flores en una espiga
interrumpida pedunculada"). Linneo escribió la palabras cataria en el margen
del nombre apropiado de la especie, que quiere decir "relacionada con los gatos",
haciendo referencia a un atributo familiar de la especie. Tanto él como sus discípulos
adoptaron el nombre Nepeta cataria para esa planta, y rápidamente sus
contemporáneos y los legos que hacían sus propias colecciones empezaron a llamar
a las especies del libro con su nombre binomial.
(28/11/2017, 15:08, Martes.)

Linneo publicó Philosophia Botanica dos años antes que la Flora
global en la que introduciría la nomenclatura binomial de cada especie, Species
Plantarum. En Philosphia Botanica compara su nomenclatura con otras en
uso en ese momento, e introduce los conceptos taxonómicos que asociará a las
categorías de género, especie y variedad. La mayoría de los géneros son heredados de
autores anteriores eminentes en cada taxón, convenientemente citados al principio del
libro junto con sus obras y las bibliotecas donde podían ser encontrados, y la
nomenclatura utilizada para esa categoría es muchas veces el nombre del género tal
como lo utilizó la autoridad preferida por Linneo para ese género -si es necesario
latinizado o sinplificado-, seguido del autor del concepto taxonómico (que no
necesariamente era el autor de la primera descripción). Muchos nombres de género
fueron utilizados ya en obras de la antigua Grecia y ya se había perdido el autor
original, la descripción original o la etimología del nombre. En las últimas décadas
cuáles eran los conceptos de "grupos naturales" que debían corresponderse con los
rangos de especie, genero y familia habían empezado a consensuarse, por ejemplo,
en 1694 Joseph Pitton de Tournefort proveyó guías para describir grupos "naturales"
de plantas en la categoría de género: los caracteres debían ser reconocibles en todos los
miembros del género y ser visibles a simple vista, sin necesidad de un microscopio.
En lo posible, debían ser tomados los caracteres de la flor y el fruto, menos
lábiles que los relacionados con el hábito. Linneo creía que los grupos que
ubicaba en las categorías de especie y género, y algunas familias (grupos de géneros),
existían en la naturaleza, eran "grupos naturales", mientras que las categorías
más altas sólo fueron utilizadas por conveniencia, formando los grupos según algún
carácter visible a modo de clave de identificación. Esta distinción entre un
concepto taxonómico "natural" y uno "artificial" no era menor, ya que el creer que
los géneros fueran reales (es decir, que existen independientemente de nuestro
discernimiento), guía al investigador de una forma diferente de la que lo hace
una categoría artificial. En palabras de Linneo:
Los caracteres no hacen al género, el género nos da sus caracteres.
-Carlos Linneo (1751:119)
Con estas palabras, Linneo reconocía que las especies se agrupaban de
forma diferente si se elegían caracteres diferentes para diferenciarlas, pero había
"algo más" que la mera similitud de caracteres que agrupara a las especies en un
mismo género, y había que saber elegir los caracteres a considerar para que la
agrupación resultara correponderse con un "grupo natural". Esta afirmación, fruto
de la intuición de Linneo, no estaría asociada a un mecanismo hasta que Darwin
pudo publicar su teoría de la evolución en 1859.
Linneo agrupó a los géneros naturales en órdenes y clases artificiales,
utilizados cual clave de identificación, útiles para que los géneros sean fáciles
de encontrar en una lista o un herbario. Los caracteres que utilizón para ello fueron
el número de estambres y el número de estigmas de los carpelos, que eran estables
dentro de cada género y fáciles de contar. Por ejemplo en la clase Pentandria,
orden Pentandria Digyna, ubicó a los géneros cuyas flores albergaban 5 "maridos"
y 2 "esposas", quedando agrupadas en el mismo orden las umbilíferas como Daucus
(la zanahoria), y la ulmácea Ulmus (el olmo); a las criptógamas, como no
tienen flores, las agrupó entre las "plantas de nupcias clandestinas".
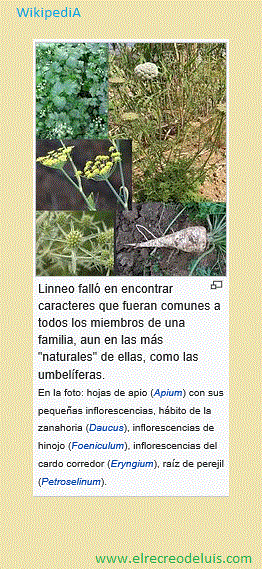
Linneo también trató de reconocer algunas familias "naturales", una
categoría intermedia entre el género natural y el orden artificial de Linneo (la
idea de que algunos grupos naturales podían ser asociados a la categoría de familia
data de Pierre Magnol en 1689), si bien falló en encontrar caracteres únicos que
pudieran funcionar como diagnósticos de las mismas, y para 1751 ya había reconocido
67 familias (Linneo 1751), dejando muchos géneros sin ubicación. Linneo buscaba
caracteres que fueran encontrados en todos los miembros de cada familia, pero
su búsqueda falló aún en las más "naturales" de ellas, como las umbelíferas.
El éxito del libro de Linneo se debía no sólo a la claridad de sus
descripciones y clasificación sino también a que se había convertido en la
revisión más exhaustiva de la literatura taxonómica hasta el momento, con una
notación cuidadosa de sinónimos. De esta forma se lo consideraba un "pasaje en
limpio" y nuevo punto de partida de la literatura taxonómica, solventando así los
problemas de comunicación producidos por la variedad de nombres y sistemas de
clasidficación locales. A partir de ese momento las actualizaciones de la
clasificación hechas por el mismo Linneo o por autores posteriores, intentaron no
abandonar la nomenclatura binomial ni el sistema de clasificación impuestos por
Species Plantarum.

En 1758 le llegaría el turno a los animales cuando Linneo publica la
décima edición de su Systema Naturae, en el que por primera vez los clasificó
como había hecho en 1753 con las plantas, adoptando la nomenclatura binomial. Linneo
nombró unas 4.400 especies de animales, incluyendo Homo sapiens, lo que
demuestra que la época no tenía reticencias en clasificar al hombre con los
animales. Su concepto taxonómico de especie, sus "tipos", eran los que hoy llamamos
ecotipos, con diferenciación de nicho (la "esencia" de sus caracteres, o atributos,
ya que los de Lineo incluían el factor genotípico y el fenotípico) y localización
geográfica, una delimitación diferente de la de la comunidad reproductiva que
hibrida en simpatría (lo que posteriormente Mayr llamaría "la especie biológica"),
a veces más atomizada y a veces más agregada que la última.
Lineo en Philosophia Botanica (1751) definió el rango de variedad,
pero no en la definición con la que hoy lo incluimos en el sistema, no era en palabras
modernas un "rango lineano". Metodológicamente, el concepto taxonómico de variedad
era el conjunto de plantas tomadas de nicho original de la especie que era criado
en condiciones diferentes, obteniendo después de una o pocas generaciones diferentes
fenotipo a partir del genotipo original. Las plantas mantenidas en el nicho del
ecotipo no pertenecían a ninguna variedad.

En las especies lineanas, el genotipo es invariable, y se encuentra en
cada una de sus variedades.
Las excepciones fueron siendo agregadas al sistema de clasificación,
autores subsecuentes clasificaron algunas especies en un orden diferente que el
resto del género. Desde la aparición de los libros de Linneo a varios autores,
como a Michel Adanson (1763-1764 citado en Judd e al. 2007), se les hizo evidente
que todos los caracteres podían variar y que no hay caracteres "esenciales" para
definir a un "grupo natural", que debían ser definidos sólo mediante un conjunto
de caracteres tomados de todo el organismo ................
(30/11/2017, 15:53, Jueves.)
........... , de los cuales la mayoría, pero no necesariamente todos, estarían
en cada organismo de la especies, en cada especie del género, en cada género de la
familia. Así se abandonaba el concepto de taxón monotético al tiempo que
se empezaba a entrar en una era de taxonomía cuantitativa, o politética, con
numerosos caracteres combinados en un diagnóstico.
En 1778 Lamarck sugirió que los caracteres utilizados por los científicos
para dividir los taxones no necesariamente tenían que ser los mismos que la gente
utilizaba para identificar algún organismo encontrado. Lamarck promovió separar
la clasificación de las claves de identificación, clasificaciones artifiales en las
que los usuarios no expertos podían llegar al taxón natural de un organismo dado
mediante caracteres que estuvieran a su alcance y con reglas de decisión explícitas.
De este forma se mitigó el conflicto entre los creadores de los sistemas de clasificación,
que a medida que aumentaba la información desponible creaban clasificaciones cada
vez más crípticas, y los usuarios de esos sistemas, que estaban más interesado en la
utilidad del sistema que en la "naturalidad" de sus grupos. En 1789 el exitoso tratamiento
taxonómico de Antoine-Laurente de Jussieu, Genera plantarum, ubicaría por
primera vez por un autor reconocido todos los taxones por arriba de género en
conceptos taxonómicos "naturales", iniciando la nomenclatura de las familias de plantas.
En término "Taxonomía" fue propuesto por Augustin Pyrame de Candolle
en 1813 a partir del griego ταξις (taxis, arreglo
o clasificación) y νόμος (nomos, ley), para
referir a la ciencia de la clasificación de los organismos. El término "Sistemática"
ya se había utilizado p. ej. por Lineo, y proviene del término griego latinizado
systema. No sería hasta 1940 en que el término "la Nueva Sistemática"
("the New Systematics", Huxley 1940 citado en Mayr et al. 1953) diferenciaría el
foco de atención de estos taxónomos de los que continuaban realizando una taxonomía
atribuida a la escuela de Lineo de diferenciar "tipos" y debatir la nomenclatura,
a los que seguían llamando "taxónomos" o "la Vieja Sistemática" (the Old Systematics).
Un cierto número de sistemáticos, especialmente en Francia y Alemania,
adoptó el concepto de "espécimen tipo". Estos "especímenes tipo" eran plantas
despositadas en un herbario que representaban la forma más común del taxón, o a
veces la forma que el autor consideraba la más "perfecta", de la que el resto del
taxón serían plantas degradadas.
La falta de estabilidad nomenclatural en las publicaciones taxonómicas
en las décadas siguientes a las obras de Linneo hizo peligrar su logro. Eso llevó
al debate y la formación de diferentes Códigos de Nomenclatura, que no eran
adoptados por un número significativo de taxónomos. Dos de ellos en particular
fueron, en 1813, Théorie Elémentaire de la Botanique ("Teoría Elemental de la
Botánica") donde se reglamentaba la elección del nombre válido para cada taxón de
plantas: y en 1842, bajo el auspicio de la British Association for the Advancemente
of Science, el Strickland Code (el "Código de Strickland", Stricklande et
al. 1842). Éstos serían los que, algo modificados, unas décadas más tarde se
consensuarían como Internacionales. (Faltan los cambios en relación a Philosophia
Botanica de Lineo).
De esta forma, desde Lineo hasta los Códigos, nacieron las reglas de
nomenclatura zoológica y botánica.
Taxonomía y evolución
Darwin en 1859 no fue el primer proponente, pero nunca antes la evolución
como hecho había tenido tal penetración en el ambiente científico. Cuando irrumpio
a mediados del siglo XIX su teoría de la evolución pronto se admitió, tal como
formuló el propio Darwin en El origen de las especies, que el grado de
similitud morfológica entre las especies era explicado por la existencia de un árbol
filogenético que las conectaba por parentesco de sus ramas ancestrales. Las
circunscripciones lineanas asumían que en los conceptos taxonómicos "naturales"
de género y especie no existían las variaciones de los caracteres "esenciales".
El conflicto entre teoría y colecciones se resolvería cuando se interpretaría que
todos los caracteres tienen el potencial de evolucionar dentro de un grupo,
y al inicio de la evolución de un grupo nuevo
no hay una diferenciación morfológica que justifique la creación de un nuevo taxón.
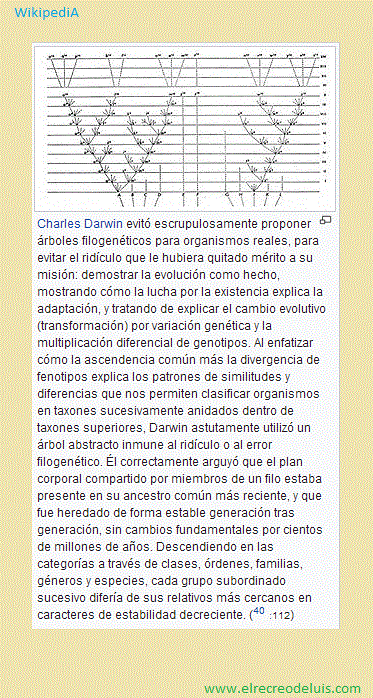
Un árbol filogenético puede reconstruirse con métodos que involucren
técnicas de agrupamiento jerárquicas, no solapadas, como son los grupos que se
usan en las clasificaciones "lineanas" (Sokal 1986), por lo que un sistema de
clasdificación lineano con conceptos taxonómicos "naturales" en principio sería,
tal como afirmaba el propio Darwin, congruente con la filogenia de las especies:
las relaciones entre taxones en las taxonomícas como indicadoras de la estabilidad
evolutiva de los caracteres que los definen. Si haber descubierto todavía la estrucura
del ADN, portador del código genético, que ocurriría recién en
los '1960 con Watson y Crick, se acumulaban las pruebas de la herencia de los
caracteres probablemente alojados en los cromosomas, para las que los redescubiertos
radios de Mendel fueron pioneros, que derivarían en la "Nueva Síntesis" a principios de
siglo XX. En pleno auge de la Nueva Síntesis, Ernst Mayr propuso fijar uno de los
conceptos de especie que hasta ese momento habían sido llamados la especie biológica
-la propuesta por Mayr, para otros autores más neutralmente se llamaría la especie
reproductiva- como el grupo de poblaciones capaz de hibridar entre sí y dar descendencia
fértil, cuyo pool o acervo genético se mentiene aislado de los de
otras en simpatría. E. Mayr suele ser citado como un taxónomo que definió el concepto
de especie que más fácilmente las delimitaba en vertebrados de tamaño grande -a los
que pertenecian los taxones en los que trabajaba-, con una precisión que en esos
taxones se corresponde mejor con los demás conceptos que en plantas y microbios.
George Gaylord Simpson (1961), descontento con la atemporalidad del concepto de
especie biológica de E. Mayr, redefinió a la especie como un linaje, una secuencia
de ancestros y descendientes con el potencial de fijar un cambio evolutivo en un carácter
(lo que Hennig llamaría una apomorfía). Si las especies se definen como poblaciones,
o como linajes, no siempre dos organismos de dos especies diferentes se diferenciarán
en términos de sus caracteres -por ejemplo, dos poblaciones todavía muy similares
en diferentes continentes, cuyo índice de dispersión no les permitió encontrarse
por un tiempo prolongado, todavía poseerán el menos potencialmente organimos que no
puedan diferenciarse entre sí taxonómicamente más que por la localidad de la
que provienen-. Hennig (1950 en alemán, 1966 en inglés) se encontró con el mismo
problema. Asumio que los linajes, que para él también se correspondían con especies,
se podían diferencia taxonómicamente en términos de sus novedades evolutivas
adquiridas (apomorfías), por lo que no habría conflicto entre la taxonomía y el
árbol de linajes. Linneo (1751) había incluido la localidad de la que provenían
los organismos en sus conceptos de especie, y también el fenotipo con el que se
encotraban en estado silvestre. La localidad y el fenotipo no volverían
a ser incorporados a las clasificaciones.
(02/12/2017, 13:15, Sábado.)
La incorporación del concepto de evolución a los taxones supraespecíficos
fue al principio modelada de forma errática. Por ejemplo, algunas clasificaciones se
se componían de taxones basados en filogenias que todavía contenían miembros
vivientes, como los árboles de Charles Edwin Bessey (1845-1915) y los de Arthur
Cronquist (1919-1992). En estos árboles no había ancestros extintos (18:43). En la
clasificación de Rolf Dahlgren, los ancestros eran sólo miembros extintos, el
árbol filogenético era un árbol tridimensional, y los miembros vivientes podían
representarse por un corte de un plano de ese árbol, en un diagrama llamado
"Dahlgrenograma" con relaciones similares a las de los países en un mapa. Un paso
crítico fue la adquisición de los métodos de reconstrucción filogenética atribuidos a
la escuela que hoy llamamos cladismo, para la cual el biólogo Wili Hennig fue el
que hizo el más valioso aporte en la década de 1960. Hennig modeló que las especies
se conectaban entre ellas a través de especies ancestrales -en su modelo original
siempre extintas-, y que podía rastrearse la trasnformación de especies ancestrales a
especies derivadas a través de las apomorfías generadas durante una cladogénesis:
los cambios de carácter en cada línea filética luego de la divergencia de un linaje
en dos. Estos árboles, hoy llamados cladogramas, fueron propuestos por Hennig como
material para construir los taxones, que siempre debían ser, a su criterio, monofiléticos.
Estos cladogramas tenían el no trivial supuesto de que la historia evolutiva de un
grupo siempre ocurre por procesos de cladogénesis (divergencia en dos y adquisición
de apomorfías). En la misma década la escuela fenética, para la que el libro de
RRSokal y PHA Sneath (1963) fue fundacional, creaba métodos numéricos de análisis
de datos intentando que fueran "ciegos", sin hipótesis de evolución previas (sin
hipótesis previas de cuáles serían las apomorfías y las plesiomorfías). Por la
época en que Hennig se popularizaba y en respuesta al cladismo la escuela tradicional,
que incorporó los métodos de construcción de dendrogramas Hennigianos y los métodos
numéricos de Sokal y Sneath en la búsqueda del árbol filogenético, pero difería
en cuál es el dendrograma que debe trasladarse a una clasificación, fue llamada
evolucionismo, la escuela evolucionista propone, como antiguamente, que puede
haber grupos parafiléticos "naturales" que deben formar parte de una clasificación
formal. El primero en utilizar el término "cladismo" en reemplazo del confuso
término Hennigiano "sistemática filogenética" fue E Mayr (1965), pero ya se utilizaba
"clado" en un sentido más amplio, según Hennig era la hipótises de ser todos los
derivados de una secuencia de ancestros y descendientes con un ancestro común, es
decir una hipótesis de grupo monofilético, otros métodos de reconstrucción filogenética
también encontrarían "clados" sin determinar cuáles eran las apomorfías y las
plesiomorfías. Hennig también redefinió monofilético, que hasta ese momento
se asumía incluyendo los grupos monofiléticos y parafiléticos sensu Hennig.
Hacia fines de la década de 1970, cladistas como G Nelson, NL Platnick
y C Patterson evocucionaron el pensamiento cladista hacia un "cladismo de patrones"
o pattem-cladism o transformed cladistics, el cual separaba teóricamente
los árboles filogenéticos, es decir los árboles de secuencias de organismos ancestros
y descendientes, de los árboles de secuencias de organismos ancestros y descendientes,
de los árboles de apomorfías, que pueden haberse correspondido o no linaje por linaje
con el árbol de organismos.

Formalización de las normas: los Códigos
El que originalmente fuera Elémentaire de la Botanique en 1813
fue reemplazo en 1930 por el altamente consensuado International Code of
Botanical Nomenclature (Código internacional de Nomenclatura Botánica, CINB),
(por sus siglas en inglés, ICBN), que trata sobre la nomenclatura de plantas y
hongos. El que originalmente fuera el Strickland Code (1842), que reglaba
los nombres de los animales, fue revisado y renombrado como International Code
of Zoological Nomenclature (Código Internacional de Nomenclatura Zoológica CINZ),
(por sus siglas en inglés, ICZN), en 1901. El Principio básico de los dos Códigos
era el Principio de Prioridad, que promovería su objetivo básico, su principio más
básico: la estabilidad en la nomenclatura. El de Botánica estableció el año de la
publicación de Species Plantarum, 1753, como el año en que se inicia la
nomenclatura botánica moderna, dejando sin efecto los nombres publicados para las
plantas antes de esa fecha. El de Zoología establecio el año 1758, el año en que
Lineo publicó la décima edición del Systema Naturae y el primero en que
utilizó su nomenclatura binomial para los animales, como el de inicio de la
nomenclatura zoológica, dejando sin efecto los nombres publicados para animales
antes de esa fecha. Los dos Códigos compartieron los mismos "principios de Nomenclatura",
y desde su aparición se realizaron periódicamente Congresos Internacionales para
su revisión.
Los bacteriólogos, que se habían atenido al Código Botánico, con el
tiempo desarrollaron su propio Código (Código Internacional de Nomenclatura de
Bacterias CINBa),(por sus siglas en inglés, ICNB) publicaco en 1958), el único
de la taxonomía biológica que posee un registro central desde 1980. El Código
Internacional de Plantas Cultivadas a su vez escindió los cultivares de las
variedades botánicas en 1952, con su última publicación en 2009, también posee
una serie de Registros de Cultivares y sus nombres son invariables, es decir
el "tipo" es todo el taxón.
Historia reciente
Desde los '1990 en que empezó a ser "tecnológicamente correcto" realizar
análisis moleculares de ADN, la sistemática en sus análisis agrega una cantidad
de caracteres de líneas de evidencia diferentes que pueden considerarse bastante
independientes entre sí: los caracteres del ADN y ARN, otras moléculas como
isozimas, flavonoides..., y caracteres de ultraestructura y micromorfología tomados
con los microscopios electrónicos de transmisión y de barrido, además de los avances
en paleontología. Estas matrices con una cantidad enorme de datos pueden analizarse
con los métodos de análisis filogenéticos para los que la escuela fenética fue
pionera, aprovechando el uso de computadoras, que han producido en las
décadas del '90 y 2000 árboles nunca antes tan consensuados y muchos de ellos hasta
inesperados, lo cual ha obligado a modificar las clasificaciones al uso, deshaciendo
grupos de larga tradición para definir otros nuevos.
Desde los '1990 que los editores publican con preferencia sistemas de
clasificación "cladistas" en colaboración de una larga lista de autores: para las
angiospermas el grupo que se ha llamado APG (1998, 2003, 2009), para los taxones
superiores Adl et al. (2005), etc.
Entre los taxónomos, todavía perdura el debate de si deben aceptarse
grupos parafiléticos o no en las clasificaciones. Los grupos parafiléticos y
monofiléticos con el mismo ancestro están compartiendo el mismo "tipo" en los
Códigos, lo cual lleva hoy en día a una gran inestabilidad en la nomenclatura.
El PhyloCode, cuyo paper seminal puede rastrearse hasta 1990 (De Queiroz y
Gauthier 1990) y cuyo primer borrador fue publicado en 1998, propone agregar las
hipótesis cladísticas al "tipo" de los Códigos en uso para restringirlo. El PhyloCode
propone nombrar formalmente sólo los grupos cladistas, y que las categorías
taxonómicas no se formalicen.
A lo largo del tiempo fueron constantes las propuestas de armonizar,
unificar, o reemplazar los Códigos, pero ninguna lograría un alto consenso (Dall
1877, ver también Savory 1962 citados en Judd e al 2007). El BioCode (primer
borrador en 1997, segundo borrador en 2011) es la propuesta más reciente de
armonizarlos, y desde 1997, se han visto unos pocos cambios en los Códigos en
uso en esa dirección. En una versión que se leía sobre el BioCode en el 2015,
éste proponía agregar más especímenes de referencia al "tipo" para restringirlo.
Otras propuestas están relacionadas con el uso de las nuevas tecnologías informáticas
y de redes para publicar nombres y crear un registro central, y sobre la eliminación
de las homonimas y la armonización de la nomenclatura de protistas.
Relación con las taxonomías populares
Sabemos muy poco acerca de la relación entre los naturalistas profesionales
y los aficionados y el público en general, si bien estas relaciones forman parte
del trasfondo histórico que ayudó a modelar la biología sistemática (Judd et al.
2007, pero ver por ejemplo Allen 1976 allí citado), Sabemos que a medida que aumentaba
el número de caracteres con los que se construían las clasificaciones, los taxónomos
profesionales en las centurias subsiguientes a la publicación de los libros de Lineo
empezaron a tratar despectivamente la taxonomía lineana que todavía era la preferida
por el lego, a la que consideraban "una diversión para mujeres y niños".
Como la sistemática tuvo su origen en Europa, los primeros grupos que se
formaron fueron aquellos que eran obvios en la flora europea. Además, la disciplina
estuvo dominada por europeos durante centurias. Fuera de Europa, especialmente en
aquellos lugares que fueran colonias europeas, la dominación europea fue especialmente
evidente en que las floras de los países fueron escritas por botánicos europeos
y basadas en material depositado en instituciones europeas (Judd et al 2007:41).
Fuera de Europa, los científicos muchas veces desvalorizaron los conocimientos que
tenían sobre las especies los nativos que poseían un profundo conocimiento de la
Naturaleza en el lugar del que eran originarios, como sucedió con los pueblos
originarios americanos.

Pocas veces los conocimientos locales han sido utilizados con criterios
antropológicos o para la investigación farmacéutica. La comparación de las taxonomías
tradicionales con las científicas ha sido objeto de algunas investigaciones.

Perspectivas actuales de la ciencia de la Taxonomía
En las décadas de 1990 y 2000 se ha vuelto a poner de moda la ciencia
taxonómica en el ambiente científico, debido en parte al revolucionario enfoque
posibilitado por la cantidad de datos derivados del análisis del ADN a los problemas
taxonómicos, y en parte a la conciencia de su utilidad, dada la crisis de biodiversidad
que estamos viviendo. Nuevas tecnologías relacionadas con la digitalización de los
datos y la utilización de internet ofrecen un potencial sin precedentes para la
diseminación de los productos de la investigación, pero al mismo tiempo que éstas
emergen se observan "vigorosos intentos" hacia el encapsulamiento y el establecimiento
de nuevas barreras para la comunicación. Las nuevas herramientas disponibles generan
un debate acerca de la utilidad de las reglas de la nomenclatura en su estado
actual, y plantean la necesidad de reformar los Códigos, reformas que aún se están
debatiendo y algunas de ellas ya se han incorporado.
El impedimento taxonómico
Quizás sorprenda que la descripción de las especies hoy se calcula realizada
sólo en un 1% o hasta un 15%, porque las plantas y los animales grandes de regiones
bien pobladas como las regiones templadas están relativamente bien descriptos, pero fuera
de ellas, como en los estratos arbóreos más altos de las selvas tropicales o en
el mundo microbiano con sus tiempos generacionales tan pequeños que lo vuelven vasto
y lo hacen evolucionar a una velocidad mucho mayor que acostrubramos a vernos
rodeados, las descripciones tienen actualmente una resolución muy baja. Como muchos
autores nos recuerdan, las descripciones de especies "nunca están completas", ya
que los descubrimientos de nuevos caracteres y nuevas poblaciones que trae cada
nueva línea de evidencia y cada nueva colecta obligan a retestear las hipótesis y
chequear cómo evolucionaron los linajes y cómo se dispersaron desde la última colecta.
Las demás ciencias de la vida (life sciences) se ven limitadas cuando las
hipótesis taxonómica de las que hacen uso son poco resueltas o confiables, lo que
vuelve los objetivos de esta ciencia hoy en día muy alejados de la demanda, a lo
que se refiere desde la Convention on Biological Diversity de 1995 como
"el impedimento taxonómico". Las causas del impedimento fueron llamadas
por los científicos "la crisis taxonómica", o también "el impedimento a la taxonomía",
en referencia a la relativa falta de recursos trasladados a afa-taxonomía a los
impedimentos sociales para su desarrollo, una ciencia que debe competir por los recursos
limitadas en universidades y subsidios con las mismas ciencias que la tienen de
impedimento.
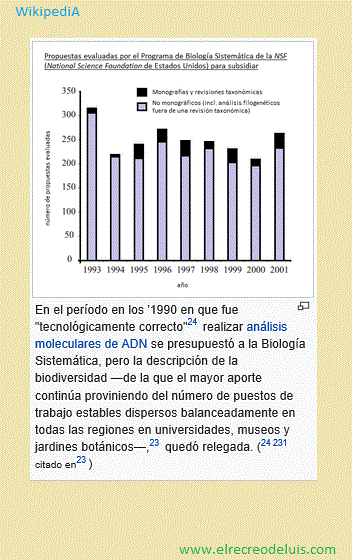
Los taxónomos descriptivos tienen pocos gastos además del salario, los
que necesitan para publicar, para realizar algún viaje y quizás algún equipamiento
adicional, de lo que lo más oneroso es el "molecular lab" del que basta con
uno cada 10 laboratorios. A lo que los taxónomos se refieren por recursos en
alfa-taxonomía es presupuestar más cantidad de taxónomos con subsidios, plazos y
evaluaciones curriculares más realistas, y el mantenimiento a largo plazo de las
colecciones:
» (1) Las colecciones a las que hoy se suman las de archivos
digitales y las de cultivos vivos de bacterias, protozoos, algas y hongos, que se
sugiere que se depositen en instituciones dedicadas como museos y jardines botánicos.
Algunos de ellos poseen colecciones ya centenarias que preservan los ejemplares
de la distribución de las especies en otras épocas y son una referencia de su
evolución, y son independientes de los arbitrios de un investigador o "pequeño lab"
que quizás se deshaga de las colecciones después de la muerte del especialista,
o quizás él mismo deba deshacerse periódicamente de las colcecciones que no utiliza
para hacer lugar en el espacio limitado que le es asignado en la universidad para
depositar colectas nuevas. Hoy se estima que se encuentran depositadas entre 1,2 y 2,1
o 3 mil millones de unidades. Las colecciones de lo que se ocupan en parte sus
curadores mediante los intercambios con otras colecciones, quienes también hacen
el valioso trabajo de reidentificación de especies para garantizar su actualidad
y evitar propagaciones de errores, y
» (2) El entrenamiento y salario de nuevos taxónomos quienes,
con los datos crudos tomados de sus colectas y otras previamente despositadas
en colecciones, publicar trabajos de literatura taxonómica primaria (sólo preparados,
la finalidad del impresionante programa PEET, no funcionó porque luego no consiguieron
trabajo.)
Como la mayor parte del trabajo científico, el trabajo de alfa-taxónomo
es conducido en proyectos relativamente pequeños, con un investigador principal
con un laboratorio asignado en la universidad y dedicación part-time (en español,
"medio tiempo" quizas "media jornada" o "jornada partida") al proyecto,
y quizás dos o tres estudiantes de doctorado que ya se han ganado esa primera beca
utilizando el laboratorio de otro o investigadores part-time del staff (en
español "personal o empleado"). En taxonomía descriptiva los proyectos son particularmente
pequeños, normalmente se hace referencia a un solo investigador, "el taxónomo
especializado en tal taxón" en la región dada, que colecta y diferencia especies
en solitario con su salario cobrado en la universidad o el museo. La mayor parte
de los subsidios (grants), (en español "subsidio, conceder, asentir, otorgar, donar,
ceder") otorgados por agencias gubernamentales o internacionales dedicadas a la
manutención de la ciencia son otorgados a la "pequeña ciencia" (small science)
(en español, "pequeña ciencia"), con una duración promedio de 3 años, los investigadores
mantienen su estabilidad laboral consiguiendo empleo en una universidad -o museos
de historia natural, jardines botánicos-, a los que dedican la otra parte de su
tiempo de trabajo. En ciencia, pocos proyectos son ambiciosos e involucran homogéneos
y en cantidad gigantesca organizados alrededor de alguna instrumentación cara y dedicada,
La llamada "big science" (en español, "gran ciencia") que aporta "big data" ( en
español "muchos datos","datos importantes"), de la que los ejemplos que suelen
mencionar son la astronomía y la física de las partículas de alta energía. Las
universidades perciben un porcentaje de los subsidios adquiridos por sus investigadores
y del prestigio que éstos acarrean por lo que tienen preferencia por ocupar investigadores
que los obtengan. Las agencias de subsidios obligan a la alfa-taxonmomía a competir
con otros proyectos para ganar los (subsidios), pero ésta termina relegada por
razones en apareiencia emocionales -muchos usuarios y hasta científicos ven a la
alfa-taxonomía o taxonomía descriptiva como una "ciencia polvorienta"-, y no pareciera
que los desperdigados taxones de los que se ocupa su piso pudiera afectar el
trabajo del resto de la facultad.
(03/12/2017, 18:04, Domingo.)
Los investigadores que ya tienen su propio laboratorio son los que eligen
de entre los estudiantes de sus materias a los que pedirán un subsidio y un puesto
de trabajo y que potencialmente competirán con el mismo investigador que les está
dando una oportunidad, esto ha sido mencionado como impedimento, (en rta. a237)
pero no hay diferencias en ese punto entre las demás ramas de la ciencia y la
alfa-taxonomía (233-238-248). El mecanismo para evaluar el trabajo de un científico
tradicionalmente es su visibilidad, de esa forma se promociona la calidad y se desalientan
las conductas inadecuadas. Pero hace décadas que se lo evalúa en lo que se
convirtió uno de los puntos más sobresalientes en su currículum: la cantidad de
publicaciones en revistas científicas y el prestigio de cada revista en la que se
ha publicado medido en forma de "Factor de impacto". este último calculado
por el Institute of Scientific Information (ISI), desde los '90 parte de
Thomson Reuters. El factor de impacto como actualmente calculado es, en
cada año determinado, de las revistas indexadas en la Web of Science (WoS)
-índice mantenido por Thomson Reuters-, la cantidad de veces que se han citado
durante el año los artículos publicados en esa revista en los 2 años anteriores a
ese, dividido el número total de artículos publicados por esa revista en esos dos
años. Los científicos intentan publicar en revistas de alto impacto para alimentar
su currículum, pero éstas suelen rechazar los trabajos de índole taxonómica debido
a que ocupan mucho espacio en la revista y argumentan que su baja cantidad de citas en los
2 años siguientes les disminuirá el factor de impacto. Las publicaciones taxonómicas
son largas, requieren muchas ilustraciones, y típicamente tienen una audiencia muy
especializada que dura por muchos años: la vida promedio de una monografía
taxonómica puede calcularse en décadas o hasta centurias, en lugar de en meses o
en años como el común de las publicaciones. Al autor a su vez le toma años o incluso
décadas la realización de una monografía, sus trabajos más acabados los publican
al final de su vida. Parte fundamental aunque implícita de la estructura de un trabajo
científico, artículos muy diversos utilizan por muchos años la información o las
claves recogidas de cada publicación de alfa-taxonomía, pero no hay costumbre de
citarlos en las referencias (algunas revistas requieren que al nombrar un taxón se
cite el autor del nombre y la primera descripción, pero tampoco requieren que se
los cite en las referencias). La información recogida impresiona como si no
necesitara un autor que la respalde, no hay interés de las editoriales que corren
tras la métricas de factor de impacto en citarlos, ni una vez publicados se agrega
la cita correspondiente ni los lapsos temporales adecuados en los índices de los que
se hacen los análisis de ránkigs, por lo que los autores de las descripciones y claves
quedan fuera del rango reconocido de publicaciones en lo emocional y en las métricas
de factor de impacto. La necesidad de cumplir con un umbral de impacto, que no mide
la calidad ni la necesidad de la descripción de un taxón, sino la "atención" que se
presta al mismo, también aumenta la disparidad en las descripciones taxonómicas:
ha provocado la deserción en sub-campos de investigación pequeños o impopulares, por
lo que los llamados "taxones subrepresentados", pobremente descriptos, todavía
son ignorados; en el otro extremo se le está dando una importancia desmedida, debido
a su popularidad, a los estudios que testean las hipótesis taxonómicas exclusivamente
por métodos moleculares y a las publicaciones sobre los muy publicitados ADN
barcodes, si bien ésta es sólo una de las herramientas utilizadas por los taxónomos
para testerar los límites de los taxones y para identificarlos. El número de citas
se reduce a las que hacen los pocos taxónomos especializados en el mismo taxón en
los dos años siguientes a las esporádicas publicaciones, y a su vez si estos colegas
publican en las pocas revistas taxonómicas cubiertas por el índice de Thomson Reuters.
Los trabajos publicados para el lego o no intencionados para ser citados, como las
guías de campo, no son valorados. Las monografías, floras y faunas parecen un trabajo que
se hace con un salario que se pagará de todos modos, no se premian con subsidios
los trabajos terminados. El taxónomo no percibe la conveniencia de publicar todo
el material que tiene, y una fuente de conocimiento valiosa se pierde con el retiro
del especialista, volviéndose atrás en las hipótesis taxonómicas sobre su taxón. La
discriminación de taxones se pasa de forma más segura de investigadores antiguos a
jóvenes de boca a boca durante la colecta, y siempre hay que revisar que sea verdadero
cada punto del material bibliográfico disponible. La alfa-taxonomía claramente
necesita que se amolde a su medida el modo de evaluar sus publicaciones científicas,
en esta "era del factor de impacto".

PA Lawrence nos recordaba en el 2008 que la forma más segura de evaluar
un trabajo científico es leyéndo -la forma habitual de hacerlo en el pasado-. El
problema de este enfoque es que sólo evalúa en retrospectiva, sobre trabajos que
tomaron muchos años de realización.
Algunas conductas hacia la nomenclatura formal pueden reducir la
calidad de la información taxonómica y repercutir en el prestigio de la rama en
general. No hay requisitos mínimos para describir una especie, y algunos autores
están de acuerdo en nombrar una colecta de la que no pudo determinarse el taxón al
que pertenecía luego de la revisión taxonómica siempre que esté correctamente
depositada en una colección accesible por otros taxónomos, como si se fijara un
epíteto a un "tipo" para después circunscribir las especies cuando pueda presupuestarse,
pero hay autores que denuncian que algunas colectas están siendo nombradas formalmente
sin que el paper (en español, "papel, documento, artículo, periódico, diario,
prueba, paquete, efecto." brinde la posibilidad a los demás biólogos de retomar
o testear sus hipótesis, o sin haber hecho una revisión taxonómica exhaustiva que
lo justificara. Miles de nombres nuevos se publican por año, de los cuales un
porcentaje muy alto, que según el autor y el taxón tratados puede ser estimado
entre el 25 y el 75%, serán nombres sinónimos de nombres previos de especies más
ampliamente distribuidas o más variables que lo originalmente descripto. En plantas
angiospermas, quizás el grupo más estudiado, se calcula que hay 5 veces más nombres
científicos que especies según los conceptos taxonómicos actuales. No parece que se valoren
los data papers ( en tafalles, "documentos de datos"), las publicaciones que
exponen los datos de las colectas sin cambios en las hipótesis taxonómicas -que a
pesar del nombre, incluyen muchas hipótesis taxonómicas: los caracteres, las
apomorfías, las especies según cada autor...-, en cambio los "papeles, documentos,
artículos", que contienen actos nomenclaturales como el establecimiento de nombres
nuevos parecieran asumirse como una señal honesta de la cantidad de trabajo que los
autores se habrán tomado. Los nombres no tienen que estar solamente establecidos,
tienen que ser nombres "robustos".

La autoría de la publicación de los datos primarios asociados
a las publicaciones primarias, como fotografías digitales de los caracteres,
secuencias de ADN, datos de ocurrencia o de colección y de estados de los caracteres
de cada espécimen de referencia

evaluaría al taxónomo con la evaluación de la calidad de los mismos y del
trabajo que respaldan. Los datos crudos son el fundamento del trabajo taxonómico,
pero hace muchas décadas que se bajaron los costos de la publicación en papel y a
pesar de que revistas electrónicas como Zootaxa y Phytotaxa y repositorios
externos con la infraestructura y la capacidad de almacenamiento requeridas han
reabierto esa posibilidad al 2015, como en biología en general, la situación no se
ha revertido y todavía son raramente publicados, como son raramente citados
formalmente. Cuando se les prgunta, la mayoría de los científicos contesta que guarda
datos no publicados de experimentos publicados y también de experimentos "que no
dieron" (que no rechazaban una hipótesis nula y no serían aceptados para publicación),
que "descansan en el sector inferior de la cajonera del lado izquierdo de su escritorio",
y la mayoría de las veces termina en la basura al momento del retiro del especialista,
"aún en los retiros más planificados", porque nadie -ni siquiera el científico- sabe
exactamente qué hay en esos medios de almacenamiento y formatos que van quedando
obsoletos ya que les falta la documentación adecuada para retomarlos cuando los
metadatos no anotados ya fueron olvidados. En las encuestas como la publicada en Huang
et al. (2012) o la hecha por Thomson Reuters en 2010 (Fig.1) la mayoría de los
científicos estaría de acuerdo en publicar sus datos en internet de forma open access
(en español, "acceso abierto"), si recibiera reconocimiento por ello en alguna forma
similar a la publicación en revistas científicas, como coautoría o citas indexadas en bases
de datos utilizadas en cálculos de métricas relevantes como las mediciones de
factor de impacto. Sin embargo, y a pesar de que parece evidente que la publicación
de los datos aumenta el número de citas de la publicación científica que hace uso
de ellos, al 2010 la proporción de datos subidos a GBIF, el repositorio más grande
de ocurrencias de especímenes creado en el 2001 para satisfacer un objetivo clave
de la Convention on Biological Diversity, "convención sobre la diversidad
biológica", bajo año a año en relación a la que se detecta o "encuentra", y de ella
sólo fragmentos están asociados a publicaciones científicas, siendo la mayor parte
datos de colecciones financiadas por organizaciones gubernamentales, y más de la
mitad de los que lo habían hecho que contestaron la encuesta denegaban la responsabilidad
de la calidad de los datos subidos a ese repositorio. Al 2013 está demostrado que
una política que hiciera mandataria la publicación de los datos con un aseguro de
su preservación y acceso al momento de la publicación de "papel, documento, artículo"
altera este paisaje para los autores, y desde 2011 se han reforzado políticas en esta
dirección por parte de varias editoriales, gobiernos y organizaciones (p.ej. Canadá,
USA, UK, la National Sciencie Foundation NSF, Evolution, The American Naturalist,
Journal of Evolutionary Biology, otras adherentes a la política JDAP, PLOS journals).
El mismo presidente Obama resaltó en su discurso al estilo inicio de sesiones State
of the Union 2012 que la "data managemente", 'gestión de datos', seá una de las
habilidades críticas de trabajo en este siglo XXI.
(05/12/2017, 15:16, Martes.)
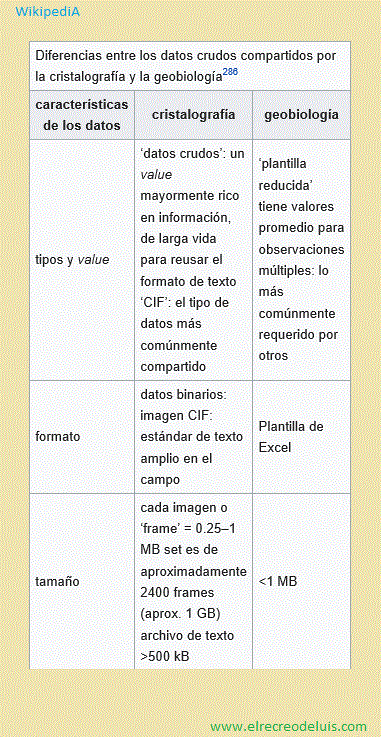
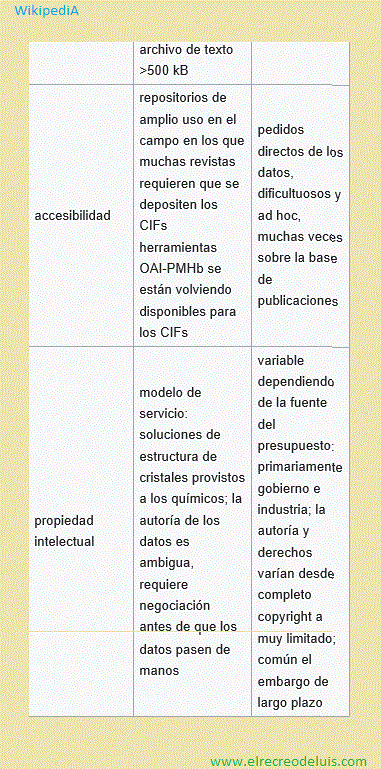
Variadas propuestas se han hecho del tipo de modernizar la infraestructura
para mejorar la eficiencia del trabajo y facilitar la evaluación del mismo, algunas
de las cuales se verán en las secciones siguientes. La que sobrevuela todas ellas
es la de digitalizar y proveer acceso online a los recursos taxonómicos. Se propone
digitalizar la literatura en papel, que las publicaciones sean de contenido libre
(ej.232,307,308, pero ver 309), que los museos digitalicen sus colecciones, que
éstas sean de contenido libre, que hubiera "megarevistas" electrónicas para las
publicaciones taxonómicas -que pueden llevar más de mil páginas-, que éstas sean
open-access, (en español, "acceso abierto"), que se alojen los datos primarios
utilizados en las publicaciones en bases de datos públicas para verificación, que
esos datos estén relacionados con claves interactivas, que éstas sean accesibles,
que sean actualizables. Son muchas las propuestas de un único portal que tenga
acceso a todas las bases de datos, tanto de colecciones como de bibliografía
(p.ej. EOL), a veces asociadas a la propuesta de ralacionar esos datos en una
única clasificación, también se propone relacionarlos mediante buscadores
estandarizados, como son comunes las propuestas de crear listas de taxones establecidos.
Tecnologías que se promueven son la digitalización 3D de imágenes de preparados al
microscopio (ej228,rSEM317), la creación de fotografías focus stacking
(en español, "apilamiento de enfoque") de especímenes tipo, ensamblarlas en
imágenes 3D, que los museos compren la tecnología para el análisis de ADN y quizás
encontrar barcodes (en español, "códigos de barras") (ej. Tautz et al. 2003),
o que haya un "molecular lab" (en español, "laboratorio molecular") cada 10
laboratorios, que se implemente la toma de datos telemétricos y monitoreo
satelital de distribución y ecología (Patterson et al. 2006). Estas iniciativas
son tecnológicamente sencillas, en este momento las principales dificultades están
relacionadas con su integración: la falta de estandarización de los formatos -para
lo cual está trabajando el TDWG-, las dificultades en la creación y mantenimineto
de registros, índices, e identificadores Globales reusables (algunos proponen en
bases de datos copiables y cosechables por buscadores específicos), y que la
financiación no debe perder el objetivo de obtener descripciones de especies publicadas,
que incluyan claves de identificación bien documentadas y accesibles que eviten
la duplicación de esfuerzos en el futuro, lo que han llamado a veces peyorativamente
"la actitud hacia el usuario final" o del usuario final (end-user attitude). Se han
financiado proyectos "horizontales" en miras de organizar una "big science" (en
español "gran ciencia") articulada, como GBIF (repositorio de ocurrencias y fichas
de colección de especímenes) y los proyectos asociados de digitalización de los
catálogos e museos y herbarios; y el registro de nombres y actos nomenclaturales
Zoobank y algún proyecto de digitalización de catálogos e de bibliotecas y hemerotecas;
y desde el punto de vista del trabajo del taxónomo, al '2010 ha demostrado en vistas
del número real de especies -mucho más alto que el estimado en centurias previas-,
organizarse por taxones de tamaño más pequeño que los Reinos gobernados por los
Códigos. Las monografías son "consumidoras de tiempo más que consumidoras de dólares"
además del salario, y los subsidios son pocos y muy grandes y corto plazo para el
equipo y el tiempo que necesita cada taxónomo, por lo que falta de taxónomos en cada taxón
podría solventarse otorgando subsidios de tamaño más realista y con evaluaciones
curriculares más realistas (PEET podía haber otorgado 10 veces más subsidios 10
veces más pequeños, a 400 Labs (en español, y español de América "laboratorios"),
en lugar de 40, por los 5 años que duraba el subsidio; la Flora of Nort America
(en español, y español de América "Flora de América del Norte" podía haber
pagado pequeños subsidios a quienes colaboraron en ella a medida que iban terminando
su trabajo), en lugar de ser un equivalente a "ganarse una lotería". En cada taxón
de tamaño más pequeños cuando cada cuerpo mandatario internacional lo decida puede
organizarse una nueva fecha de inicio de la nomenclatura, la que hoy en día
sólo existe en bacterias, fijadas en 1980.
El copyright ("derecho de copia"), (en españo y español de América
"derechos de autor"), es de concernimiento en la preservación de la literatura
taxonómica. La literatura taxonómica primaria en una distribución sin formato no
se vería afectada por las leyes que restringen la realización de copias y su
distribución (p.ej. el Convenio de Berna), pero son pocos los investigadores que
publican comentarios al respecto (p.ej. Costello 2009) y parece que las editoriales
no se hacen eco del tema. Unicamente las publicaciones de De Carvalho y colaboradores
brasileños denuncian que los "piratas de la biodiversidad" pueden boicotear
el trabajo del taxónomo.
El rol del ADN y los "marcadores de ADN"
Los análisis moleculares de ADN se utilizan hoy habitualmente en los
análisis filogenéticos y desde los '90 que unos pocos cientos de bases, con su
cantidad enorme de combinaciones potenciales, parecen bastar para hacer análisis
de identificación y parentesco. Esto llevó a algunos autores, como Hebert et al.
(2003) y Tautz et al. (2003), a proponer asignar un rol central al ADN en la
descripción e identificación de las especies, de forma que una muestra de ADN y la
lectura de una de sus secuenciaa de bases debería ser una especie de marcador para
ubicar el taxón al cual pertenece un espécimen, de una forma similar a como se
usa el código de barras en los supermecardos. Tautz et al. incluso proponen reformar
los Códigos para que este "identificador" sea un carácter agregado al "tipo". Esta
"Taxonomía basada en ADN" aún adolecería de muchos de los mismos problemas que
tienen las demás propuestas de identidicaciones o de agregar atributos al "tipo",
en particular, hay que decidir qué secuencia usar, ya que algunas secuencias no dan
información que diferencie al taxón de los demás. Esto puede ser porque un mismo gen
puede mantenerse inalterado durante millones de generaciones y muchas especiaciones
después de su formación, o debido al fenómeno de introgresión, de forma que un gen que
se había diferenciado vuelve a su estado anterior. Por lo tanto, de la misma forma
en que no es conveniente confiar en un solo carácter morfológico para identificar
una especie, tampoco es conveniente confiar en una sola secuencia de ADN. Además,
aun cuando la "Taxonomía basada en ADN" fuera financiada, habría que preguntarse
si es necesario agregar un requerimiento extra al ya lento proceso de describir
nuevos taxones, en especial teniendo en cuenta que se calcula que sólo el 10% de las
especies del planeta ha sido descrito. Debido a eso, probablemente la mayoría de los
biólogos verá las secuencias de ADN como un complemento de la información morfológica,
más que intentar reemplazarla buscando verdaderos barcodes (en español y
español de América "códigos de barras"). De todas formas, los Códigos no especifican
ningún carácter en particular para diagnosticar nuevos taxones, así que la
"Taxonomía de ADN", con el tipo nomenclatural de costumbre, ya es válida, aunque
la identificación mediante caracteres visibles puede ser de uso más inmediato y su
realización más económica que la lectura de un verdadero barcode (en español
y español de los americanos "código de barras").
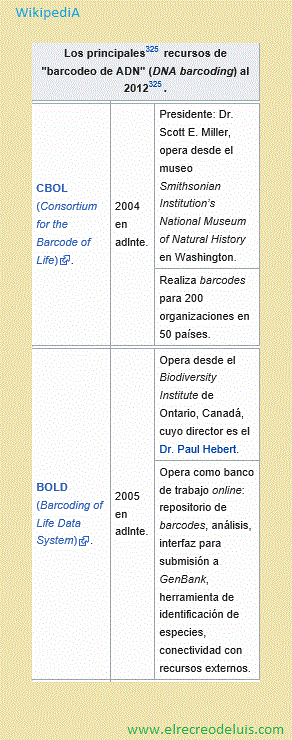
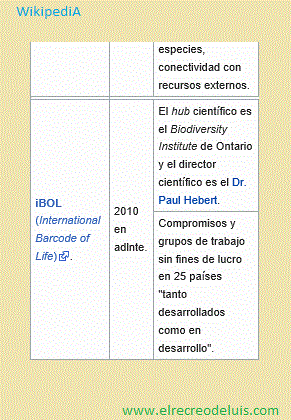
Al 2003 ya era rutina que los microorganismos se delimitaran con métodos
moleculares, y está claro que para dilucidar el árbol de la vida completo, sería
útil secuenciar los mismos genes en muchos taxones diferentes. Para lograr esto, sería
necesario un "proyecto genoma horizontal" y un sistema de archivo de secuencias
de ADN como el INSDC que incluye al estadounidnese GenBank, (en español y
español de los americanos "banco de genes"), más allá de si el ADN se vuelve un
requerimiento en la descripción o identificación de especies o si se lo agrega
al "tipo". .....................
ensp;(06/12/2017, 12:50, Miércoles.)
........... Al 2005, dos proyectos habían recibido el millonario presupuesto
para llevar a cabo la empresa global de secuenciación de barcodes
(en español y español de los americanos "código de barras") -nombre que quedó a las
secuencias seleccionadas por los proyectos presupuestados-, el Barcode of life
Data System (BOLD), (en español y español de los americanos "Sistema de datos de
la vida en código de barras"), y el Consortium for the Barcode of Life (CBOL,
"Consorcio para el código de barras de la vida"). Sus seguidores promocionan los
casos de macroorganismos en que el agregado de esa línea de evidencia molecular a
las hipótesis taxonómicas llevó al esclarecimiento de problemas de sinonimias, como
cuando la secuencia de ADN "barcode", (en español y español de los americanos
"código de barras") COI logró diferenciar en 4 especies "crípticas" la sanguijuela
medicinal europea. Como en ese mismo ejemplo, la secuencia estandarizada de ADN
que es parte del gen COI tomada de un espécimen es de utilidad para delimitar
especies sólo acompañada de la matriz de caracteres del espécimen tomados de otras
líneas de evidencia, moleculares y no moleculares, y de una preidentificación morfológica
-QD Wheeler nos recuerda que muchos especímenes de referencia descansan en las colecciones
sin una caracterización morfológica adecuada, y en un porcentaje significativo está
mal la determinación o no se ha intentado su identificación-, sólo de esta forma puede
utilizarse esta secuencia en una matriz de análisis filogenéticos, y posteriormente
quizás forme parte de una clave de identicicación. A diferencia de lo que ocurrió
en los '90 en que los "barcodes" (en español y español de los americanos
"código de barras") se tomaban sólo sobre epecímenes morfológicamente caracterizados
hasta hipótesis de especies, al 2013 los barcodes (en español y español de
los americanos "código de barras") se han producido a una velocidad mayor que la
revisión taxonómica que debería acompañarlos, lo que ha hecho proliferar en el
estadounidense GenBank (en español y español de los americanos "banco de genes")
los llamados taxones "en la oscuridad" (dark taxa). Éstos podrían formar parte de
taxones conocidos en los que el organismo del que se extrajo el ADN aún no haya
sido ubicado, o de taxones nuevos, no caracterizados. Muchos barcodes
(en español y español de los americanos "código de barras"), "sin nombre" (unnamed)
habían sido suprimidos por GenBank (en español y español de los americanos
"banco de genes"), pero aun así su número siguió aumentando en los últimos años.


En respuesta a la intensa propaganda realizada a favor de esta empresa
-que para muchos autores desbalanceaba las necesidades presupuestarias de la
alfa-taxonomia-, el término "taxonomía integrativa" fue acuñado, independientemente
y casi al mismo tiempo, por Dayrat (2005) y Will et al. (2005), el último luego de
una conferencia, el DNA barcoding debate (en español y español de los
americanos "Debate de códigos de barras de ADN") Will vs. Hebert, el "padre de
barcode". La taxonomía, de integrar líneas de evidencia durante la generación
de hipótesis taxonómicas, y en particular a la integración de los variados métodos
moleculares con los morfológicos como la única primera aproximación válida para
delimitar taxones en la categoría especie. Desde esa fecha una considerable número
de estudios taxonómicos se hizo eco del término.
La presupuestación de los proyectos promovidos por Hebert et al. (2003)
dejó como legado lo que algunos autores llaman la primera región de ADN "horizontal"
estandarizada para su utilización como atributo en una matriz de análisis filogenéticos,
para lo que anteriormente se habían utilizado muchas regiones de ADN nucleares y
organelares diferentes enfocadas en taxones de tamaño más pequeño. (ver p.ej. en
plantas). QD Wheeler decía en el 2004: "Estamos en el umbral del salto más excitante
de la 'Taxonomía basadas en morfología' [identificación de especies mediante claves
morfológicas] desde Linneo (1758), lo que vuelve estos tiempos un momento inoportuno
para darse por vencido". ..."Los datos visuales morfológicos nunca habían podido ser
eficientemente comunicados en el pasado. Ahora tenemos la tecnología para una
edad dorada de la morfología comparativa si podemos sobrellevar las presiones
para conformarnos a lo 'tecnológicamente correcto'".
La crisis de biodiversidad
La crisis de biodiversidad causada por actividades humanas, como las que
llevan a la pérdida de hábitat y la introducción de especies exóticas, es tal vez
comparable en magnitud a las grandes extinciones registradas en los períodos geológicos
(PH Raven 2002). El término "biodiversidad" fue acuñado por EO Wilson como un
reemplazo eufónico de "diversidad biológica" en una serie de populares ensayos
donde desde mediados de los '1980 ponía el foco en ésta y su crisis; en 1992, el
tratado internacional llamado Convenio sobre la Diversidad Biológica,
(Convention on biological Diversity), ratificado por todos los países miembros
de la ONU salvo Estados Unidos, reconoció como un derecho internacional la valoración
de la biodiversidad en el contexto de su conservación y uso sustentable. Sin embargo,
existe una conciencia general de que no será posible conservar toda la biodiversidad.
Los especialistas deben calcular en qué hábitats y en qué especies focalizar los
magros presupuesto asignados a la conservación, priorizar los hotspots
(en español y español de los americanos "puntos calientes")
(regiones con una concetración de especies endémicas con peligro de perder el
hábitast), y generar modelos predictivos acerca del futuro de la biodiversidad.
A 1995, se había hecho evidente para el CBED que los especialistas no pueden medir
una biodiversidad no descripta, lo que bautizaron como "el impedimento taxnómico",
a raíz del cual el organismo impulsó el desarrollo de la alfa-taxonomía. Las medidas
concretas encaminadas a cumplir estos objetivos empezaron a ser visibles en algunos
países e instituciones internacionales a fines de 1990, momento a partir del cual
se hizo evidente un segundo impedimento: la elección del concepto taxonómico a
utilizar tien sus consecuencias enlas decisiones de conservación. Por ejemplo,
Peterson y Navarro-Sigüenza (1999) analizaron la cantidad de especies de aves
endémicas de Mexico utilizando dos clasificaciones diferentes basadas en
distintos conceptos de especie. Según el concepto biológico de especie o BSC
(que agrupa dentro de la misma especie a los organismos con capacidad de
hibridar y dar descendencia fértil), obtuvieron 101 especies, con la mayor
concentración de especies endémicas en las regiones montañosas del sur y del oeste.
En cambio según el concepto filogenéico de especie o PSC (que asigna como
miembros de una especie a los que poseen evidencia de conservar caracteres en un
análisis de filogenia, más allá de si pueden hibridar con otras poblaciones o no),
obtuvieron 249 especies endémicas, con la mayor concetración en zonas tanto llamas
como montañosas del oeste. La elección de una clasificación de cada hábitat. La
estabilidad que presentaron por al menos dos décadas desde las influentes
publicaciones de E Mayr donde proponía su concepto de especie biológica hasta -1985
algunos taxones bien conocidos, contrasta fuertemente con la reciente incertidumbre
sobre el concepto de especie a utilizar y la "inflación" en su número, que
especialmente en los grupos bien estudiados, incluso en primates, ocurrió en
parte significativa por un cambio en la preferencia de los taxónomos del concepto
de especie biológica o BSC al concepto de especie filogenética o PSC.
La preferencia de conservación de la categoría especie trajo un problema
extra a la clasificación. Esto ha animado la elevación al rango de especie a las
poblaciones que necesitaban protección, más allá de si la decisión podía ser
respaldada científicamente. El fenómeno fue descrito en la tortuga negra
Chelonia agassizi, pero es probable que haya ocurrido también en otros
taxones. Esta inflación de especies estaría sesgada hacia formas carismáticas,
de gran tamaño, raras y en peligro de extinción, que atraigan interés de público
y científico para su conservación. Un interés local puede llevar a una población
a ser listada aparte, pero su sobrevivencia puede ser más probable si se la
gestiona como un componente de una metapobalción viable. (Isaac et al. 2004).
Ha habido otras propuestas de medidas de biodiversidad, con sus
consecuencias en las propuestas de conservación, se ha propuesto como medida de
biodiversidad la categoría de género (incluso en primates, como en Harcourt et
al. 2002) o las poblaciones, otros estudios abandonaron la taxonomía biológica
para describir la biodiversidad con medidas de "diversidad filogenética". Sin
embargo, esas medidas pueden quitar atractivo presupuestario debido a que la
opinión pública favorece los taxones especie como uidad de medida. Una solución
sería que los checklists o inventarios (listas de especies por región)
se agreguen de forma globalizada especificando el concepto de especie utilizado
y se enlazaran a sinonimias bajo otros conceptos de especie, de forma que se
sincronicen todas las clasificaciones que afectan una región para presentar su
diversidad taxonómica sin dejar de lado ninguna autoridad. La tecnología para
hacelro está disponible, pero se necesita una inversión institucional fuerte y
de largo plazo.
Informática de la Biodiversidad
El campo de acción de la informática de la biodiversidad (biodiversity
informatics (en español y español de los americanos "informática de la biodiversidad")
son las ciencias biológicas que utilizan como unidad de trabajo los organismos,
lo que la diferencia de la ecoinformática (ecoinformatics, la informática
al nivel de organización de los ecosistemas) y la bioinformática (bioinformatics,
al nivel de organización molecular y fisiológica).
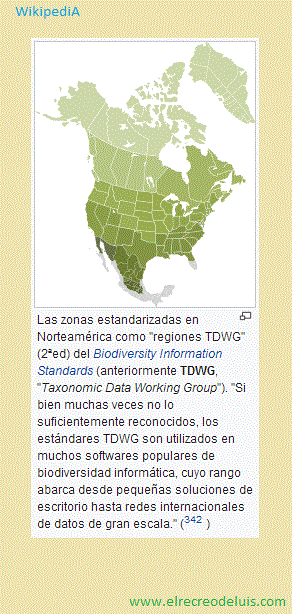
El material tomado como digital o a digitalizar en taxonomía consta de
los "datos crudos" y sus metadatos, y de la "literatura taxonómica primaria" y
sus respectivos metadatos. La digitalización requiere un id "único" pra cada uno
de ellos, idealmente "global", es decir único en internet. Por ejemplo la forma
más directa de crear un id global es concatenar el id del dato con el de su tabla
en la base de datos con el identificador de esa base de datos en intenet. Hay
varias propuestas de formación de id globales y alguna de ellas puede volverse
estandarizada en el futuro, cuando la aceptación por parte de la comunidad de
usuarios lo decida. En internet además se busca que este GUI (identificador global
único, global unique identifier (en español y español de los americanos
"identificador único global") pueda "resolverse", enlazarse a través de internet
a un archivo con sus metadatos y quizás una lista de enlaces a copias del dato
taxonómico, los dos alojados en su base de datos online (en español y
español de los americanos "en línea"), en lo que cuando son adecuademente citados
se llamó el mundo emergente de los Linked Open Data (datos abiertos
vinculados), de La Big New Biology (nueva gran biología), la "Nueva
Biología Gigante" más "data-céntrica". Las discusiones en la comunidad de la
informática de la biodiversidad se enfocan en 3 tipos de GUIs "autorresolubles,
es decir que sean una dirección de internet que al tipearla en la barra de
direcciones devuelva la página con los datos o los metadatos: los HTTP URIs
(es decir URLs), los GUI recomendados como estándar por el TDWG que son los LSID
(que hacen uso de los DNS de internet, y resuelven a una página de metadatos),
y los DOI, que hacen uso de una autoridad central para resolverse y son por
ejemplo los asignados por CrosssRef para publicaciones científicas que los resuelve
al dato. Los GUIs no ocupan espacio en las publicaciones taxonómicas porque los
metadatos y el material al que hacen referencia pueden agregarse en bases de
datos externas, las bases de datos deberían ser "copiables" para amortiguar la
posibilidad de que metadatos y datos se pierdan, y para independizarse de las
autoridades que mantiene las bases de datos mediante buscadores -sarch engines-
(en español y español de los americanos "motores de búsqueda") específicos o
inespecíficos que indexen los GUI, sus metadatos, sus locaciones, sus GUI
sinómimos y usos posteriores de los mismso. Como la unidad de trabajo en la
clasificación es el organismo, los datos crudos y la literatura primaria deberían
enlazarse a los id de los organismos o cepas refenciados en ellos, por lo que
implican la necesidad de un GUI por organismo/cepa. Los id de los organismos son
el taxón terminal de las clasificaciones según cada autor. Cada nombre de taxón
sensu cada fuente también necesita un id, para poder separar sinónimos
nomenclaturales del nombre y sincronizar conceptos sinónimos con diferente nombre.
Los datos primarios y su literatura asociada pueden agregarse en proyectos
"verticales" de un taxón algo superior o con líneas de evidencia que requieren
diferentes especialistas, la digitalización de los metadatos primarios que tengan
"actos nomenclaturales" (establecimientos de nombres, etc.) por taxón "vertical"
en miras de crear las tan esperadas Listas de Nombres Adoptados (no suprimidos)
necesarias para los Códigos en uso es condiserada una prioridad para el BioCode,
(en español y español de los americanos "biocódigo"). También hay proyectos
"horizontales" como los de ADN barcodes = codibarras, la Biodiversity
Heritage Library (en español y español de los americanos "Biblioteca del
patrimonio biodiverso") que digitaliza literatura taxonómica, los proyectos que
digitalizan los especímenes y sus metadatos (ficha de colección, etc.) de las
colecciones de los museos. Algunos sitios web, como la Encyclopedia of life
(EoL) (en español y español de los americanos "enciclopedia de la vida"), son
"iniciativas globales", funcionan como portales que presentan la información
"agregada" de "asociados" (partners) (en español y español de los americanos
"compañero, socio, asociado, copartícipe, pareja, interlocutor, cónyuge,
firmante, acompañar, estar asociado con"), las bases de datos indexadas en ellos.
El candidato obvio para ser el id que combinara bases de datos diversas había
sido el nombre científico, como fue utilizado en iSpecies y EOL, pero pronto se
vieron sus limitaciones como identificadores debido a que no son ni completamente
estables ni globalmente únicos: en los nombres científicos son permitidas, o
inevitables, las sinonimias y las homonimias. Idealmente en el futuro los nombres
científicos sean reemplazados por id de conceptos taxonómicos (de nombres sensu
cada autor), que puedan resolverse hacia la cita a la que corresponde la descripción
del taxón utilizada con todas sus referencias.
(07/12/2017, 21:04, Viernes.
Entre las iniaciativas más populares desde el 2003 quizás estén:
» GBIF Financiado por agencias gubernamentales en acuerdos internacionales
luego del CBD. Portal de acceso a los datos de ocurrencias provenientes de cientos
de instituciones de todo el mundo. Recomienda enviar las colaboraciones individuales
a iNaturalis, eBird o la Encyclopedia of Life (EOL).
» Species 2000. Al 2015 se lee: "ITIS y Species 2000 se han asociado
para formar el Catálogo de la Vida (Catalogue of Life, Col) que orgullosamente
provee el esqueleto de la Enciclopedia de la Vida (Encyclopedia of Life)".
La Enciclopeda de la Vida nació en el año 2003 y puede agregarse información luego
de registrarse como usuario. Es muy publicitada en Science (por ejemplo en
Stuart et al. 2010).
» Tree of Life, desarrollado en los '90 y anunciado formalmente en 1996,
el esqueleto taxonómico lo provee la comunidad científica, la comunidad de usuarios
puede agregar contenido multimedia por taxón. Al 2007 se lee, "muchos usuarios" y
contribuyentes de Tree of Life (Árbol de la Vida) han notado el extenso
solapamiento en los objetivos de EOL y ToL, y ha habido algún concernimiento
acerca de la supervivencia a largo plazo de ToL. En orden de evitar la duplicación
de esfuerzos, ToL y EOL han acordado coordinar cercanamente las actividades futuras,
compartir contenido y herramientas de "compuintangibles". El proyecto se dejó de
actualizar en el 2011.
Pero hubo más de 50 proyectos en todo el mundo. Un problema común a muchos
de ellos fueron las expectativas irrealistas de los usuarios respecto al número de
datos que debían ser curados, por lo que luego de la fundación inicial los recursos
se desvanecían y los proyectos quedaban detenidos. Un ejemplo fue All Species
Foundation (Fundación de todas las especies), creado en el 2001 por un ambicioso
grupo de "Silicon-Valley types" enre los que estaban prominentes investigadores como
EO Wilson de Harvard, y Peter Raven del Missouri Botanical Garden. (Jardín botánico
de Misuri). Su objetivo era catalogar todas las especies del planeta en 25 años, pero
en el 2005 el proyecto había muerto. Se habían terminado las donaciones luego de
finalizada la "explosión de la burbuja punto com" (dot.com bubble burst), una visita
del 2005 a su página web sólo decía, "Nos fuimos" ("Gone"). Ni los assets (activo)
fueron reutilizados. EO Wilson ya había retomado la idea en su nuevo proyecto, la
Encyclopedia of Life (EOL) (Enciclopedia de la vida, EDLV), con el objetivo
a plazos más modestos de una página web para cada especie del planeta, presentada
por el mismo EO Wilson en el 2003.
Pero si bien ya en el 2010 Padial et al.229, calcularon que tomaría 400
años de trabajo taxonómico constante llegar a describir las cerca de 10 millones de
especies de eucariotas, al 2012, la realidad es que las colecciones de historia
natural están casi tan lejos de la digitalización completa como 20 años atrás, y
aún no hay una verdadera database central de acceso a colecciones, ni un registro
completo de las mismas para ofrecer a los usuarios. A través de GBIF, la fuente
más grande de información, Ariño (2010) estimó que eran accesibles no más del 3%
de los datos de especímenes preservados en las colecciones de todo el mundo.
Bladoderov et al. (2012) calcularon que el costo de la digitalización de los cerca
de 30 millones de especímenes de insectos alojados en la colección de entomología
del Museo de Historia Natural de Londres, por ejemplo, tomaría 23 años de trabajo
continuo de las 65 personas que componen el staff del departamento, y en el mundo,
dependiendo de la colección los costos varían entre 0,50 a unos cuantos dólares por
espécimen para capturar sus datos completamente, por lo que la digitalización completa
de todas las colecciones de historia natural del planeta costaría unos 150 mil
millones de euros y tomaría unos 1500 años. La solución más común propuesta es la
prioridad de la digitalización sobre la base de la demanda, también se propone
financiar los costos de una digitalización "industrial" de las colecciones.
(08/12/2017, 10:44, Viernes.)
Zootaxa fue la primera "megarevista" electrónica con referato. Creada en
el 2001, llegó a publicar monografías de más de mil páginas, y en el 2004 ya se
perfilaba como la mayor revista en taxonomía zoológica, momento en el que entró
a las mediciones de factor de impacto de ISI. Al 2008 se promocionó como la
primera en publicar un "paper cibertaxonómico" (artículo cibertaxonómico), con
enlaces y otras utilidades. A ella siguió una generación de nuevas "revistas rápidas"
en taxonomía, incluyendo Phytotaxa (2009) y Zookeys (2008). Pensoft
(Zookeys, Phytokeys, Mycokeys) distribuye desde el 2011 de rutina los datos
taxonómicos publicados hacia una serie de recursos online (en línea) que incluyen GBIF,
EOL, y la indexación en Zoobank, al 2015, una actualización del 'compuintangible'
permite que estas bases de datos externas "cosechen" los datos de los papers (documentos,
artículos) que corrigen datos de papers (documentos, artículos) anteriores, así los datos
se corrigen dinámicamente en
ellas. Estas revistas también hicieron las primeras pruebas de publicar papers
(artículos, documentos) "de datos" (data papers), (documentos de datos).
En el 2012 los dos Códigos permitieron la publicación electrónica de actos nomenclaturales,
facilitando el uso de este recurso.
Algunas revistas tradicionales hoy exigen que los datos crudos (raw data)
(datos sin procesar) sobre los que fueron tomados los datos de publicaciones
taxonómicas sean alojados en bases de datos públicas. También son usuales los
artículos "suplementarios" publicados en formato exclusivamente electrónico.
Por otro lado, al 2014, las bases de datos externas ofrecen la posibilidad
de publicar monografías taxonómicas fuera del circuito de las revistas científicas,
con las facilidades que podría dar una base de datos interactiva. Algunos
'compuintangibles' para monografías son las plataformas Scratchpads, KE-EMU,
Diversity Workbench Platform (plataforma de trabajo del banco de la diversidad)
EDIT Platform for Cybertaxonomy (plataforma para la cyber taxonomía) (el
último el único de los listados con capacidad de alojar clasificaciones alternativas),
'compuintangibles' para alojar una base de datos descriptivos y crear claves son
Lucid2, Xper2.
La armonización de los Códigos y el BioCode
La IUBS es un organismo que tiene su participación en los Códigos en uso
y en 1985 formó un Comité de Nomenclatura Biológica, el cual desde ese momento ha
organizado numerosos congresos y reuniones, a los que se han sumado el auspicio
de la IUMS (International Union of Microbiological Societies)(Unión internacional
de sociedades microbiológicas) y del IAPT, y el apoyo de organismos como CAB
International (internacional), el Linnean Society of London (Sociedad
de Linnean 'Linneana'
de Londres), La Royal Society of London, (real sociedad de Londres) entre otras.
En 1994 se formó el IUBS/IUMS International Committee on Bionomenclature (ICB)
(Comité internacional de bionomenclatura). Fue el ICB quien propuso un primer borrador
de un Código unificado de nomenclatura después de sus esperables congresos y revisiones
que fue presentado como el "Borrador del BioCódigo 1997" (Draft BioCode 1997) (DL
Hawksworth, presidente de la IUBS).
El Biocode propuso que fuera utilizado en reemplazo de los Códigos en uso,
pero su uso no fue implementado por los autores de publicaciones taxonómicas. La
crítica principal consistía en que había baja disposición de botánicos y zoólogos
en aprenderse todo un conjunto de reglas nuevas, argumentando que los problemas
existentes podían superarse reformando cada Código en uso, lo que quizás está
relacionado con la crítica de haber sido "poco maduros" al presentar una propuesta
semejante.
La crítica fue escuchada y la nueva versión del BioCode, el Draft
BioCode 2011 (Borrador del BioCódigo 2011), ha sido sustancialmente reescrita de
forma no de reemplazar los Códigos actuales, sino para proveer un marco común que
los englobe. Está repensado para ser utilizado junto con los Códigos en uso -a los
que llama los Códigos Especiales-, como su complemento, y está listo para su implementación
inmediata. Es un "terso texto de sólo 35 artículos", que refiere a los demás Códigos
cuando lo considera apropiado, pero para la mayoría de los propósitos puede funcionar
por sí mismo. Propone un número de mejoras futuras, que dependen de que se hagan
reformas en los Códigos Especiales para poder efectuarse "y que esperablemente
disparen esas reformas". Una de ellas es el requerimiento de registrar nombres
nuevos y actos nomenclaturales (por ejemplo las designaciones de lectotipos), que
en a nueva versión fue modificada para tomar ventaja de los recientes progresos
tecnológicos. Otras innovaciones propuestas son el requerimiento de que las descripciones
de nuevos taxones sean en inglés o latín (en el 2011 todavía se debían hacer en
latín para plantas, y en cualquier lengua en animales), la opción de proteger nombres
y sus atributos por medio de Listas de Nombres Adoptados, el prospecto de evitar
los homónimos entre-Códigos para los nombres futuros, y el permiso, para los llamados
organismos "ambirregnos", de usar sufijos de cualquier Código para los nombres
supragenéricos, independientemente del Código que se esté aplicando. Introduce
rangos adicionales a los primarios (profamilia entre familia y subfamilia, progénero
entre género y subgénero, proespecie entre especie y subespecie) de forma de
implementar las reglas de forma retroactiva para todos los tipos de organismos sin
efectos desestabilizadores en el sistema actual de nombres. (W Greuter, secretario
de la ICB).
(09/12/2017, 11:51, Sábado.)
A comienzos de este siglo, la rápida evolución de las bases de datos
dejó en evidencia que es más práctica la producción de listas de nombres adoptados
por taxón, y la nueva edición fue creada para que sus provisiones puedan ser
adoptadas en diferente momento por diferentes taxones dentro de cada Código, lo
cual estaría determinado por el cuerpo internacional mandatario apropiado para cada
taxón si fuera operacional (DL Hawksworth)
Algunos cambios menores fueron realizados en los Códigos Especiales desde
1997. Con la llegada de los análisis moleculares aumentó la cantidad de grupos que
fueron tratados sucesivamente bajo los Códigos de Botánica y Zoología, por lo que
se acentuó la necesidad de armonizar la nomenclatura de protistas, que al 2011 queda
pendiente. También queda pendiente un registro mandatario de nombres y una lista
de nombres en uso, que al 2011 sólo sigue existiendo en bacterias (DL Hawksworh)
Las variaciones de terminología entre Códigos, y la terminología unificada
propuesta:
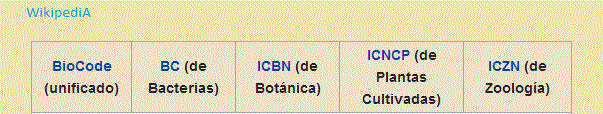
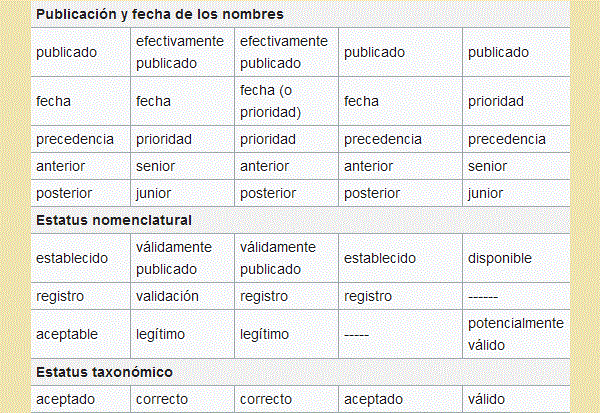

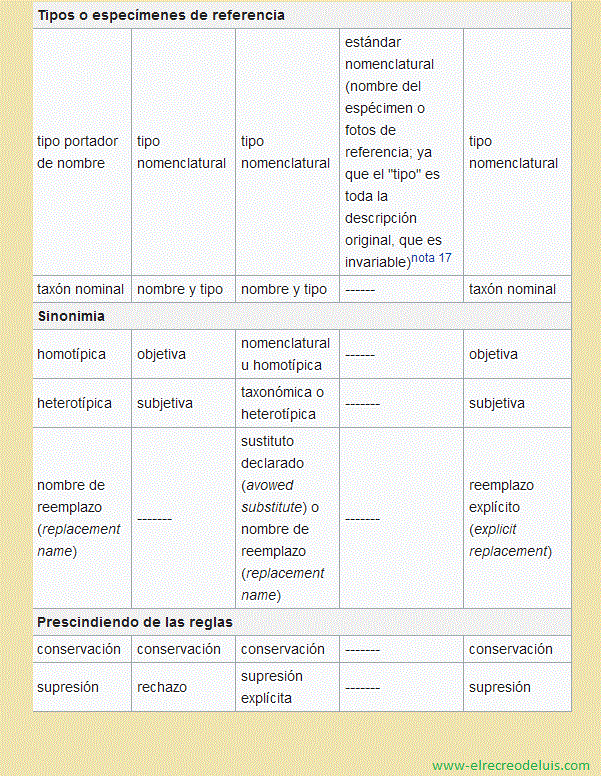
Propuestas de un registro central de nombres
Para cerrar las "listas de nombres adoptados" Charles Godfray (2002)
propuso una "primera revisión a través de la web" como el nuevo punto de partida
de la nomenclatura en cada grupo, de forma que luego de una exposición por un
tiempo predeterminado de las listas provisorias con su revisión taxonómica abierta
a los agregados de información nueva por parte de la comunidad con acceso a internet,
éstas fueran cerradas y las descripciones previas encontradas posteriormente fueran
consideradas como descripciones con nombres informales. Propuestas como ésta suelen
estar acompañadas por la propuesta de registrar los nuevos actos nomenclaturales
en alguna database central (base central de datos), de forma que el nombre no fuera
establecido si no fue registrado. Alessandro Minelli (2003), el presidente de la
Comisión Internacional para la Nomenclatura Zoológica, también argumentó sobre lo
prioritario de contar con un registro central de nombres. El Código de Bacterias
ahora son publicados en una única revista, con una nueva fecha de comienzo de la
nomenclatura fijada en 1980. Se esperaban reformas similares en los Códigos de
macroorganismos, pero zoólogos y botánicos todavía tienen que ratificar las propuestas.
La posibilidad de que unos pocos especialistas pudieran dejar publicaciones fuera
de la indexación y monopolizar la nomenclatura de un grupo había sido una de las
preocupaciones que llevó a los botánicos a rechazar las llamadas para un registro
central de nombres en el Congreso Internacional de Botánica en 1999, propuestas
similares fueron rechazadas en el Congreso de Zoología de 1999.
La posible reglamentación que había sido parte del borrador del BioCode
1997, fue modificada en el borrador del BioCode 2011 para incorporar las nuevas
tecnologías y la posibilidad de dividir los taxones abarcados por los Códigos en
grupos de tamaño más pequeño, cada uno con su propio cuerpo mandatario internacional.
En el año 2012 una enmienda del Código de Zoología oficializó un registro central
de nombres llamado Zoobank, la enmienda no modifica sin embargo la fecha de inicio
de la nomenclatura de cada grupo, ni el registro es mandatario, sólo insta a que
a partir del momento se informen las publicaciones taxonómicas en esa web. En
Botánica sólo el Mycobank, que había sido creado en el 2004, se adoptó como
registro mandatario desde el 2013. Al 2015 las listas no están cerradas.
El PhyloCode
Algunos autores armentan problemas usando el sistema linneano de clasificación
cuando se lo basa en el árbol filogenético (p.ej. de Queiroz y Gauthier 1990). El
PhiloCode (http://www.ohio.edu/phylocode), creado en 1998 y actualizado periódicamente,
está basado en lo que sus autores llaman una "aproximación filogenética" a la
nomenclatura. Con el objetivo de ser un código de nomenclatura, tiene "varias
similitudes en sus metas y en sus métodos" con lo que los autores llaman la
"nomenclatura basada en rangos" (lo que otros autores llaman la nomenclatura
linneana): provee métodos inambiguos para aplicar nombres a los taxones, para seleccionar
un único nombre aceptado para cada taxón, y no impide a los taxónomos aplicarles
rangos taxonómicos, "contrariamente a un concepto erróneo frecuente". Una de las
diferencias es que el PhyloCode aplica los nombres a "la secuencia de ancestros
y descendientes, o sus productos, los clados"; en lugar de aplicarlos "a una
circunscripción + rango". De esta forma el Código define un taxón nominal cuando
realiza la asociación entre un nombre y un clado, o más rigurosamente, entre un
nombre y las hipótesis cladísticas delimitadas por los especificadores (otros
taxones o especímenes de referencia y apomorfías) de un clado, como publicadas en
la primera descripción; lo que los autores llaman las "definiciones del nombre".
Tres tipos de "definiciones del nombre" son la basada en nodos, la basada en ramas,
y la basada en apomorfías. Una definición del nombre Asteraceae podría ser
"el clado que se deriva del ancestro común más reciente de Barnadesia y Aster".
Los nombres de los taxones no cambiarían con los cambios de circunscripción mientras
se mantenga ese tipo, y como esos nombres no dependen del rango taxonómico (como
en la nomenclatura de los Códigos que tiene reglas relacionadas al respecto, como
el uso de sufijos), entonces cuando hay cambios de categorías taxonómicas no es
necesario que cambien los nombres de los taxones (de Queiroz 2012).
Los autores del PhyloCode pretenden reemplazar los Códigos de la nomenclatura
linneana con su nuevo Código, lo cual no llegó a discutirse en el debate acerca de
una taxonomía unitaria en el ambiente científico.
Sin embargo, el PhyloCode ha sido intensamente debatido en revistas
cieníficas de prestigio como Taxon, y hasta aparecieron remeras universitarias
con inscripciones "anti-rangos" que fueron sospechadas de propaganda hacia ese
Código. La crítica principal hacia el PhyloCode es que el sistema lineano es igual
de compatible con el árbol filogenético, por lo que no hay necesidad de cambiar
todo un sistema nomenclatural por otro. De hecho el PhyloCode puede no ser compatible
con el árbol filogenético en la categoría especie, ya que no maneja todos los conceptos
taxonómicos utilizados en esa categoría. Otra crítica es que ese "tipo" no probó
ser en realidad más estable, "a menos que se redefina estabilidad", por lo que aun
dentro de una "filosofía taxonómica" cladista sería difícil evitar la convivencia
con otros Códigos especialmente en categorías muy altas. Quitar las categorías
absolutas quitaría información sobre exclusiones y la mayoría de los taxónomos
ve como suficientemente útil para comunicación que estén impresas en los sufijos
como para abandonarlos en las categorías del grupo familia. (pero ver) La posibilidad
de que más de un espécimen de referencia, y las apomorfías o la descripción de un
epíteto descriptivo puedan agregarse formalmente al "tipo" -lo último de hecho
mandatario en taxones de categorías más altas-, y otras propuestas se podrían hacer
bajo los Códigos en uso (p.ej. 381,109), que serían aprobadas después de un debate
adecuado y un amplio consenso. Los "nombres descriptivos" de categorías altas de
hecho cuando hacen referencia a un carácter o un plan corporal sobresaliente se
interpretan como derivados de hipótesis cladísticas que contienen las apomorfías
que definieron el plan corporal en su linaje ancestral y que le dan el nombre
al grupo -que no necesariamente es monofilético-. Por ejemplo Cavalier-Smith (2013)
define al reino Fungi como derivado del linaje que adquirió rizoides absorbentes,
tempranamente acoplado en su desarrollo a la adquisición de una pared celular de
quitina con pérdida de la fagocitosis, poropone a Excavata como un grupo parafilético,
y propone abandonar el nombre del supergurpo Unikonta ("un cilio"), ya que se
había determinado que el cilio se había origindado dos veces independientemente
dentro del grupo. Del aluvión de publicaciones referentes al PhyloCode, las dos
lecturas recomendadas por el FAQ del ICZN son Wheeler (2004) y Knapp et al. (2004).
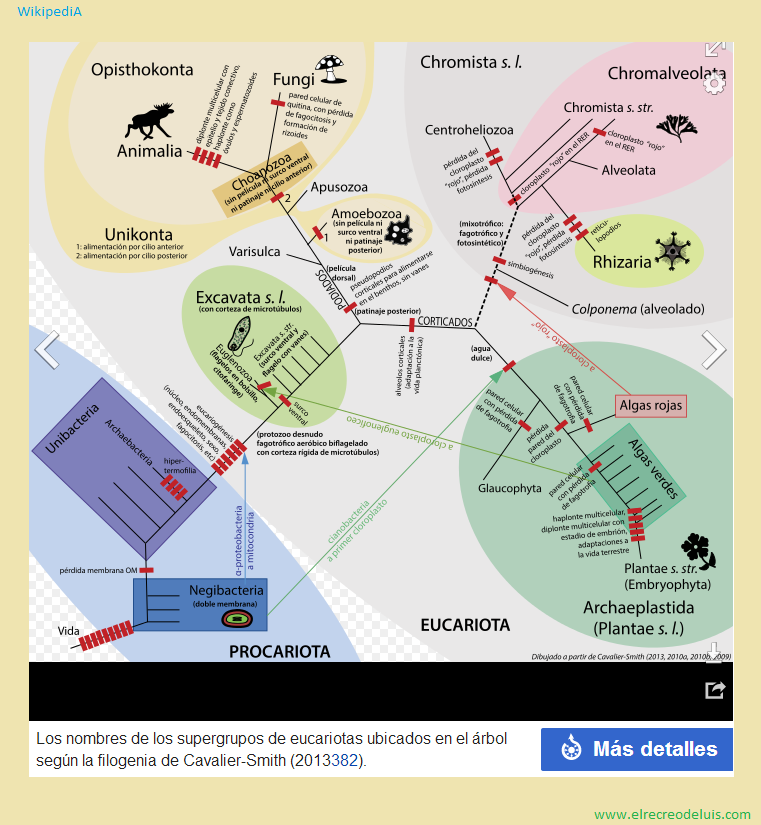
El PhyloCode fue sólo una de las manifestaciones de los taxónomos que,
en general bajo la nomenclatura de los Códigos en uso, señalan que es buena
práctica que sea explicito el "concepto de taxón" utilizado (p.ej. 120,140,125,135
para los conceptos especie, cen categorías intermedias). En taxones intermedios,
un ejemplo del 2007 en subfamilias de rosáceas ha quedado así:

La diferencia entre ambos Códigos está en el tipo, que se traspasa de
forma inalterable de taxón a taxón.
<>·Taxonomía linneana<>:
WikipediA - 15/10/2017 - 17:16 - Domingo. ;
La taxonomía de Linneo o taxonomía linneana clasificaba a los
seres vivos en diferentes niveles jerárquicos, comenzando originalmente por el de
Reino. Hoy, se considera el Dominio como una jerarquía suprarreinal,
dada la reciente necesidad de incluir también a Bacterias y a Arqueas. Los reinos
se dividen en Fios o Phyla (en singular, Phylum) para los
animales, y en Divisiones para plantas y otros organismos. Estos se dividen
en Clases, luego en Órdenes, Familias, Géneros y Especies.
Aunque el sistema de Carlos Linneo era firme, la expansión de conocimientos
ha dado lugar a una expansión del número de niveles jerárquicos, incrementando los
requerimientos administrativos del sistema, aunque permanece, es el único sistema
de clasificación básica que actualmente cuenta con la aprobación científica universal.
Entre las subdivisiones posteriores, han surgido entidades como superclases, super,
sub e infraórdenes, super y subfamilias, tribus y subtribus. Muchas de estas
jerarquías adicionales suelen surgir en el estudio de disciplinas como la entomología,
que requiere clasificar nuevas especies. Cualquier campo biológico que estudie las
especies está sujeto a la clasificación taxonómica linneana, y en extensión, a sus
rangos jerárquicos, particularmente si se lleva a cabo la integración de organismos
vivientes con especies fósiles. Será conveniente entonces aplicar herramientas
más novedosas d clasificación, como la cladista.
Tras el rango de especie, se pueden dar también subrangos, tales como
subespecie y raza en animales, y variedad y forma en botánica,
aunque, en esta última disciplina, el término subespecie también es utilizado.
<>·Teca<>:
rae; Del griego ???? th?ke 'caja'. 1. f. Cajita donde se guarda un --recuerdo--. 2. f. Bot. Célula en cuyo interior se forman las esporas de algunos hongos.
·Tegumento:
rae; Del latín ·tegumentum·, "lo que cubre, cubierta". 1. m. Biol. Órgano que
sirve de protección externa al cuerpo del hombre y de los animales, con varias capas
y anejos como glándulas, escamas, pelo y plumas. 2. m. Biol. Membrana que cubre el
cuerpo del animal o alguno de sus órganos internos.
·Telofase:
rae; Del al. Telophase, y este del gr. t???? télos 'fin, término' y el al. Phase
'fase'. 1. f. Biol. Cuarta y última etapa de la mitosis, en la que los cromosomas
se distribuyen en las dos células hijas y el citoplasma se divide en dos.
/// ·Telofase, mitosis:
WikipediA -MITOSIS- (05/11/2016); La telofase (del griego te???, que significa 'finales'),
es la reversión de los procesos que tuvieron lugar durante las profase y prometafase.
Durante la telofase, los microtúbulos no unidos a cinetocoros continúan alargándose,
estirando aún más la célula. Los cromosomas hermanos se encuentran cada uno asociado
a uno de los polos. La envoltura nuclear se reforma alrededor de ambos grupos cromosómicos,
utilizando fragmentos de la envoltura nuclear de la célula original. Ambos juegos
de cromosomas, ahora formando dos nuevos núcleos, se descondensan de nuevo en cromatina.
La carocinesis ha terminado, pero la división celular aún no está completa. Sucede
una secuencia inmediata al terminar.
·Telómero:
Rae; Del griego télos ·fin, término· y meros ·parte·. 1. Biol. Extremo de
un cromosoma.
·Temperatura:
rae; Del latín ·temperatura·. 1. f. Magnitud física que expresa el grado o
nivel de calor de los cuerpos o del ambiente, y cuya unidad en el sistema internacional
es el Kelvin (K). 2. f. coloq. Estado de calor del cuerpo humano o de los seres vivos.
<>·Temporal1<>:
R.A.E.; De latín «temporalis» derivado de «tempus», -oris 'tiempo'.
1. adj. Perteneciente o relativo al tiempo.
2. adj. Que dura por algún tiempo.
4. adj. Que pasa con el tiempo, que no es eterno.
5. adj. Gram. Que expresa tiempo. Adverbio temporal. Aplicado
a una oración.
6. m. Tempestad (\\ tormenta grande).
7. m. Tiempo de lluvia persistente.
8. m. Trabajador rústico que solo trabaja por ciertos tiempos del año.
<>·Tentativo, va<>:
R.A.E.; Del latín medieval «tentativus», y este del latín «tentatus»,
participio pasivo de «tentare» 'tantear, experimentar, e «-ivus» -ivo'.
1. ad. Que sirve para tantear o probar algo.
2. f. Acción con que se intenta, experimenta, prueba o tantea algo.
3. f. Examen previo que se hacía en algunas universidades para tantear
la capacidad y suficiencia del graduado.
4. f. Der. Principio de ejecución de un delito por actos externos que
no llegan a ser los suficientes para que se realice el hecho, sin que haya mediado
desistimiento voluntario del culpable.
<>·Teoría<>:
R.A.E.; Del griego
[θεωρια - "theoría"].
1. f. Conocimiento especulativo considerado con independencia de toda aplicación.
2. f. Serie de las leyes que sirven para relacionar determinado orden de fenómenos.
3. f. Hipótesis cuyas consecuencias se aplican a toda una ciencia o a parte muy
importante de ella.
<> · Teoría neutralista de la evolución molecular <>:
WikipediA - 14/11/2017, 15:53, Martes.;
La teoría neutralista de la evolución molecular establece que la
gran mayoría de los cambios evolutivos a nivel molecular son causados por la
deriva genética de mutantes y no mutantes neutros en cuanto a la selección natural.
La teoría fue propuesta por Motoo Kimura a finales de los 1960s y pricipios de los
1970s, y aunque fue recibida por algunos como un argumento contra la teoría de
Darwin de la evolución por medio de la selección natural, Kimura mantuvo (con el
acuerdo de la mayoría de quienes trabajan en biología evolutiva) que las dos teorías
son compatibles: "La teoría no niega el papel de la selección natural en la
determinación del curso de la evolución adaptativa". En cualquiere caso, la teoría
atribuye un gran papel a la deriva genética.
...
<>·Teórico, ca<>:
R.A.E.; Del latín tardío «theoricus», este del griego
[θεωρικος - "theorikós"].
1. adj. Perteneciente o relativo a la teoría.
2. adj. Que conoce las cosas o las considera tan solo especulativamente.
3. adj. Dicho de una persona: Que cultiva la parte teórica de una ciencia o
un arte.
5. f. p. us. Teoría (// conocimiento especulativo con independencia de toda
aplicación).
/// ·Termófilo:
WikipediA - (21/11/2016-lunes); El término termófilo se aplica a organismos
vivos que pueden soportar condiciones extremas de temperatura relativamente altas,
por encima de los 45ºC. Es un subtipo de vida extremófila. Muchos termófilos pertenecen
al dominio Archaea. Estos organismos tienen una tasa de crecimiento rápida pero de
corta duración. Ejemplos de este tipo son los organismos vivos que se desarrollan
al lado de las fumarolas o chimeneas hidrotermales en las profundidades del océano
como el anélido Riftia pachyptila y también algunos microorganismos como las
bacterias metanógenas que se desarrollan en las mismas condiciones.
·Terrestre:
rae; Del latín ·terrestris·. 1. adj. Perteneciente o relativo a la tierra.
2. adj. Terrenal. 3. adj. Perteneciente o relativo a la tierra en contraposición
al cielo y al mar.
<>·Tetra-<>:
rae; Del griego tet?a- tetra-. 1. elem. Compos. Significa 'cuatro'. Tetrasílabo, tetrápodo.
<>Tetraedro<>:
rae; Del griego
·τετραεδρον· 'tetráedron'.
1. m. Geom. Sólido terminado por cuatro planos o caras.
<>Tetraedro regular<>:
rae;
1. m. Geom. Tetraedro cuyas caras son triángulos equiláteros.
<>·Tiempo<>:
R.A.E.; De latín «tempus».
1. m. Duración de las cosas sujetas a mudanza.
2. m. Magnitud física que permite ordenar la secuencia de los sucesos,
estableciendo un pasado, un presente y un futuro, y cuya unidad en el sistema
internacional es el segundo.
3. m. Parte de la secuencia de los sucesos.
4. m. Época durante la cual vive alguien o sucede algo. En tiempo de
Trajano. En tiempo del descubrimiento de América.
5. m. Estación (\\ cada una de las cuatro partes del año).
6. m. edad (\\ tiempovivido).
7. m. edad (\\ duración de una cosa).
8. m. Oportunidad, ocasión o coyuntura de hacer algo. A su tiempo.
Ahora no es tiempo.
9. m. Espacio de tiempo disponible para la realización de algo. No
tengo tiempo.
10. m. Largo espacio de tiempo. Tiempo ha que no nos vemos.
11. m. Cada uno de los actos sucesivos en que se divide la ejecución de
algo; como las composiciones musicales.
12. m. Estado atmosférico. Hace un tiempo húmedo.
14. m. Gram. Categoría gramatical deíctica que permite localizar la
acción, el proceso o el estado denotados por un verbo a partir de su relación con
el momento del habla o con otro punto temporal. En español, el verbo presenta
flexión de tiempo, modo, número y persona.
15. m. Gram. Cada uno de los paradigmas de la conjugación verbal en
función de la categoría tiempo. Los tiempos del indicativo.
16. m. Marina. Tempestad duradera en el mar. Correr un tiempo. Aguantar
un tiempo.
17. m. Mec. Cada uno de los recorridos del pistón para completar un ciclo
en un motor de combustión interna alternativo.
18. m. Mús. Cada una de las partes de igual duración en que se divide el
compás.
·Tierra:
rae; 1. Planeta que habitamos. 2. Superficie del planeta Tierra, especialmente
la que no está ocupada por el mar. 3. Material desmenuzable de que principalmente
se compone el suelo natural.
·Típico, ca
rae; Del latín tardio ·typîcus, y este del griego t?p???? typikós.
1. adj. Característico o representativo de un tipo. 2.adj. Peculiar de un grupo, país,
región, época, etc.
·Tipo, pa:
Rae; Del latín ·typus·, y este del griego t?p?? týpos. 1. m. Modelo, ejemplar.
2. m. Símbolo representativo de algo figurado. 3. m. Clase, índole, naturaleza de las
cosas. 4. m. Ejemplo característico de una especie, de un género, etc. 8. Bot. y zool.
Cada uno de los grandes grupos taxonómicos en que se dividen los reinos animal y vegetal,
y que, a su vez, se subdividen en clases.
<>·Tisular<>:
rae; Del francés ·tissulaire·, derivado de ·tissu· 'tejido'. 1. adj. Biol. Perteneciente o relativo a los tejidos de los organismos.
·Tonelada:
Rae; De ·tonel·.
1. f. Tonelada métrica. 2. f. Marina. Tonelada de arqueo. 3. f. Medida antigua para
el arqueo de las embarcaciones, igual a ocho codos cúbicos de ribera. 5. f. Tonelería
(// conjunto de toneles) 6. f. Peso de 20 quintales.
·Tonelada de arqueo:
Rae;
1. f. Mar. Medida de capacidd equivalente a 2,83 m3.
·Tonelada de peso:
Rae;
1. f. Tonelada (// peso de 20 quintales)
·Tonelada métrica:
Rae;
1. f. Unidad de peso equivalente a 1000 kg. (Símbolo t).
·Tonelada métrica de arqueo:
Rae;
1. f. Marina. Metro cúbico.
<>·Topología<>:
R.A.E.; Del griego [τοπος] "tópos" 'lugar'
y -logía.
1. f. Mat. Rama de las matemáticas que trata especialmente de la
continuidad y de otros conceptos más generales originados de ella, como las
propiedades de las figuras con independencia de su tamaño o forma.
<>·Topología arbórea<>:
WikipediA - 02/11/2017, 15:36, Jueves.;
Una configuración en árbol o topología en árbol o
estructura en árbol es una caracterización física de un objeto y sus componentes,
que por su configuración se asemeja o recuerda a un árbol, en el sentido que sus
ramificaciones tienden a converger en un origen o raíz (por ejemplo
el llamado árbol genealógico).
Con este concepto se introduce por lo tanto las nociones de raíz y
de descendencia.
En informática es vulgarmente utilizada como topología (como estructura),
junto a otras topologías en anillo o topologías en estrella. En Programación informática,
estas topologías son utilizadas como estructuras de datos para resolver problemas
complejos donde por ejemplo debe aplicarse indexación.
Enlaces en un árbol
Por definición, una estructura en árbol está constituida por nodos, y
también por definición, un árbolo vacio (Sin nodos) es también una estructura en
árbol.
Un nodo de un árbol tiene un determinado valor o contenido, y desde el
mismo (a veces llamado nodo-padre) pueden derivar uno o varios nodos-hijo.
El grado de una estructura arbórea es el número máximo de descendientes
presentes, al recorrer cada uno de los nodos de la estructura. Si de cada nodo de
la estructura se derivan como máximo otros dos nodos, se dice entonces que se tiene
un árbol binario.
Una configuración en árbol o topología en árbol, es una tal caracterización
de un objeto y de sus componentes, de manera que su configuración se asemeja a un
árbol, en el sentido que sus ramificaciones tienden a converger en una raíz única.
Aplicaciones
En ingeniería, el concepto de árbol tiene muchas aplicaciones, pues
permite caracterizar topologías y modelos, hashing, así como diagramas varios (por
ejemplo, un diagrama organizacional o un determinado tipo de red pueden ser descritas
como una estructura arbórea).
<>·Topología de red<>:
WikipediA - 02/11/2017, 17:11, Jueves.;
La topología de red se define como el mapa físico o lógico de
una red para intercambiar datos. En otras palabras, es la forma en que está
diseñada la red, sea en el plano físico o lógico. El concepto de red puede
definirse como «conjunto de nodos interconectados». Un nodo es el punto en el
que una curva se intercepta a sí misma. Lo que un nodo es concretamente depende
del tipo de red en cuestión.
Un ejemplo claro de esto es la topología de árbol, la cual es llamada
así por su apariencia estética, por la cual puede comenzar con la inserción del
servicio de internet desde el proveedor, pasando por el router, luego por
un switch y este deriva a otro switch u otro router o
sencillamente a los hosts (estaciones de trabajo), el resultado de esto es
una red con apariencia de árbol porque desde el primer router que se tiene
se ramifica la distribución de Internet, dando lugar a la creación de nuevas
redes o subredes tanto internas como externas. Además de la topología estética,
se puede dar una topología lógica a la red y eso dependerá de lo que se necesite
en el momento.
Los componentes fundamentales de una red son el servidor, los terminales,
los dispositivos de red y el medio de comunicación.
En algunos casos, se puede usar la palabra arquitectura en un sentido
relajado para hablar a la vez de la disposición física del cableado y de cómo
el protocolo considera dicho cableado. Así, en un anillo con un concentrador
(unidad de acceso a múltiples estaciones, MAU) podemos decir que tenemos una
topología en anillo, o de que se trata de un anillo con topología en estrella.
La topología de red la determina únicamente la configuración de las
conexiones entre nodos. La distancia entre los nodos, las interconexiones físicas,
las tasas de transmisión y los tipos de señales no pertenecen a la topología de la
red, aunque pueden verse afectados por la misma.
Tipos de topologías
...
·Tóxico, ca.
Rae; Del latín ·toxîcum· "veneno", y este del griego t?????? toxikón 'veneno para
emponzoñar las flechas', 'veneno', der. de t???? tóxon 'arco'.
1. adj. Que contiene veneno o produce envenenamiento. Aplicado a una sustancia.
2. adj. Pereneciente o relativo a una sustancia tóxica.
·Traducción:
rae; Del latín ·traductio, -onis "acción de pasar de un punto a otro", ·traslado·.
1. f. Acción y efecto de traducir. 3. f. Interpretación que se da a un texto.
4. f. Biol. Etapa de la expresión genética en la que se sintetiza una proteína
a partir de la información contenida en el ARN mensajero mediante el código genético.
<>·Tragar<>:
rae; De origen incierto. 1. tr. Hacer movimientos voluntarios o involuntarios de tal modo que algo pase de la boca hacia el estómago. 3.tr. Comer vorazmente. 6. tr. coloq. Absorber, consumir, gastar algo. La obra se tragó más cemento del presupuestado. 7. tr. coloq. Arg. y Ur. Entre estudiantes, empollar (// estudiar mucho). 10. prnl. coloq. Chocar con un obstáculo. Tragarse una farola.
<>·Trans- también Tras- en algunas voces
<>:
rae; Del latín ·trans-·. 1. pref. Significa 'al otro lado de' o 'a través de'. Transalpino o trasalpino, transpirenaico o traspirenaico, traspaso, trastienda.
·Transcribir también Trascribir:
rae; Del latín ·transcribere·. En Argentina, Paraguay y Uruguay Transcripto
1. tr. copiar (-- escribir en una parte lo escrito en otra).
·Transcripción también Trascripción:
rae; Del latín transcriptio, -onis. 1. f. Acción y efecto de transcribir.
2. f. Biol. Etapa de la expresión genética en la que la secuencia de ADN es copiada
en una secuencia de ARN.
/// <>·Transducción (genética)<>:
WikipediA - (05/12/2016-Lunes); La transducción es un proceso mediante
el cual el ADN es transferido desde una bacteria a otra mediante la acción de un
virus. También se utiliza para designar el proceso mediante el cual el ADN
exógeno es introducido en una célula mediante un vector viral. Esta es una
herramienta que usualmente utilizan los biólogos moleculares para introducir en
forma controlada un gen extraño en el genoma de una célula receptora.
Cuando los bacteriófagos (virus que infectan bacterias) infectan una célula
bacteriana, su modo normal de reproducción consiste en capturar y utilizar la
maquinaria de replicación, transcripción y traducción de la célula de la bacteria
receptora para producir gran cantidad de virones, o producir partículas virales,
incluido el ADN o ARN viral y la cubierta de proteína.
/// <>·Transformación (genética)<>:
WikipediA - (05/12/2016-Lunes); En biología molecular, la transformación
es la alteración genética de una bacteria resultante de la absorción directa,
incorporación y expresión de material genético exógeno (ADN exógeno). El ADN
exógeno se encuentra en el ambiente y se introduce a través de la membrana de la
célula bacteriana. La transformación ocurre de forma natural en algunas especies
de bacterias, aunque también se puede efectuar por medios artificiales. Para que
ocurra la transformación, la bacteria debe estar en un estado de competencia, lo
que puede ocurrir como una respuesta limitada en el tiempo a las condiciones
ambientales, tales como la falta de nutrientes y una densidad celular elevada.
La transformación es uno de los tres procesos por los que el material genético
exógeno se puede introducir en una célula bacteriana. Los otros dos son la
conjugación y la transducción. A la transformación de células eucariotas se le
llama transfección.
El término transformación es también usado, de manera más general, para
describir mecanismos de transferencia de ADN o ARN en biología molecular (es
decir, teniendo en cuenta más que las consecuencias genéticas). Por ejemplo la
producción de transgénicos como maíz transgénico requiere la inserción de nueva
información genética en el genoma del maíz usando el mecanismo apropiado de
transferencia de ADN; al proceso se le llama comúnmente transformación.
El ARN también puede ser transferido en las células usando métodos similares,
pero esto no provoca normalmente cambios heredables y por lo tanto no es una
transformación real.
·Transmitir también trasmitir:
rae; Del latín ·transmittêre·. 1. tr. Trasladar, transferir. 2. tr. Dicho de
una emisora de radio o de televisión: Difundir noticias, programas de música,
espectáculos. 3. tr. Hacer llegar a alguien mensajes o noticias. 5. tr. Conducir
o ser el medio a través del cual se pasan las vibraciones o radiaciones. 6: En una
máquina, comunicar el movimiento de una pieza a otra.
·Transpiración también traspiración:
rae; 1. f. Acción y efecto de transpirar. 2. f. Bot. Salida de vapor de
agua, que se efectúa a través de las membranas de las células superficiales de las
plantas, y especialmente por los estomas.
·Transpirar también traspirar:
rae; Del latín meieval ·transpirare·; propiamente "exhalar a través de". 1.
intr. Dicho del cuerpo: Exudar a través del tegumento. 2. intr. Dicho de una cosa:
Sudar (//destilar agua a través de sus poros).
<>Transportador, ra también trasportador<>:
rae;
1. adj. Que transporta. 2. m. transportador de ángulos.
<>Transportador de ángulos<>:
rae;
1. m. Utensilio semicircular graduado que sirve para medir y trazar ángulos.
·Transportar:
rae; Del latín transportare. 1. LLevar a alguien o algo de un lugar a otro.
/// Transporte trans-membranal: El transporte trans-membranal se produce
cuando las moléculas atraviesan totalmente la membrana de la célula. la difusión
facilitada (o el transporte facilitado) es un proceso de difusión transmembranal
que se produce en todas las células eucariotas. Es además una forma de transporte
pasivo, donde las moléculas pasan a través de la bicapa de fosfolípidos, con la
ayuda de las proteínas de transporte. Estas proteínas canales pueden abrirse y
cerrarse con el voltaje del ion, proveniente de un impulso del sistema nervioso,
que pasa a través del axón.
·Trasversal también Trasversal:
rae; Del latín medieval ·transversales·. y este derivado del latín ·transversus·.
1. adj. Que se halla o se extiende atravesado de un lado a otro. 3. adj. Que se cruza
en dirección perpendicular con aquello de que se trata.
·Través:
rae; Del latín ·transversus· 'oblicuo', 'transversal'. 1. m. Inclinación o
torcimiento de una cosa hacia algún lado. 3. m. Arq. Pieza de madera en que se afirma
el pendolón de una armadura. 4. m. Marina. Dirección perpendicular a la de la quilla.
a través: 1. loc. adv. de través.
de través: 1. loc. adv. En dirección transversal.
al través de: 1. loc. prepos. a través de.
a través de: 1. loc. propos. Pasando de un lado a otro de. A través de
la celosía. A través de una gasa. 2. loc. propos. Por entre. A través de la
multitud.
3. loc. prepos. por intermedio de = 1. loc prepos. Por mediación de.
·Tribu:
rae; Del latín ·tribus·. 1. f. Cada uno de los grupos de origen familiar que
existían en algunos pueblos antiguos. Las tribus Iberas y celtas de Hispania.
2. f. Grupo social primitivo de un mismo origen, real o supuesto, cuyos miembros
suelen tener en común usos y costumbres. 3. f. coloq. Grupo de individuos con alguna
característica común, especialmente las pandillas juveniles. Las tribus urbanas.
4. f. Biol. Cada uno de los grupos taxonómicos en que se dividen muchas familias
y que, a su vez, se subdividen en géneros.
/// <>Tridimensional<>:
WikipediA (26/02/2017 - Domingo. 21:09);
En física, geometría y análisis matemático, un objeto o ente es tridimensional
si tiene tres dimensiones. Es decir, cada uno de sus puntos puede ser localizado
especificando tres números dentro de un cierto rango. Por ejemplo, anchura, altura
y profundidad.
El espacio a nuestro alrededor es tridimensional a simple vista, pero en
realidad hay más dimensiones, por lo que también puede ser considerado un espacio
tetra-dimensional si incluimos el tiempo como cuarta dimensión. La teoría de
Kaluza-Klein original postulaba un espacio-tiempo de cinco dimensiones; la teoría
de cuerdas retoma esa idea y postula según diferentes versiones que el espacio
físico podría tener 9 o 10 dimensiones.
/// <>·Triptófano<>:
WikipediA - (07/01/2017-Sábado-13:36); El triptófano (abreviado como Trp o W) Es un aminoácido esencial en la nutrición humana. Es uno de los 20 aminoácidos incluidos en el código genético (codón UGG). Se clasifica entre los aminoácidos apolares, también llamados hidrófobos. Es esencial para promover la liberación del neurotransmisor serotonina, involucrado en la regulación del sueño y el placer. Su punto isoeléctrico se ubica a pH=5.89. La falta de triptófano puede contribuir negativamente a cuadros de ansiedad, insomnio y estrés.
·Tronco, ca:
rae; Del latín ·truncus·. 1. adj. desus. Trunco, truncado, tronchado. 2. m.
Cuerpo truncado. Tronco de pirámides, de columna. 3. m. Tallo fuerte y macizo
de los árboles y arbustos. 4. m. Cuerpo humano o de cualquier animal, prescindiendo
de la cabeza y las extremidades. 5. m. Conjunto de dos o más mulas o caballos que
tiran de un carruaje. 6. m. Conducto o canal principal del que salen o al que concurren
otros menores. Tronco arterial, de chimenea.. 7. m. Ascendiente común de
dos o más ramas, líneas o familias.
·Tropismo:
rae; Del griego t??p?? trópos 'vuelta' e -ismo. 1. m. Biol. Movimiento de
orientación de un organismo sésil como respuesta a un estímulo.
<>·Trufa<>:
rae; Del occitano antiguo ·trufa· este del latín vulgar ·tufêra· y este del latín ·tuber·. 1. f. Variedad muy aromática de "Criadilla de tierra". (Definición no muy consistente y demasiado escueta que no aclara ni el género ni la especie, en conclusión no es fiable, por no ser una descripción completa). 2. f. Dulce de pasta de trufa rebozada en cacao en polvo o en varillas de chocolate. 3. f. Pasta hecha con chocolate sin refinar y mantequilla. 4. f. embuste (//mentira).
·Tubo:
rae; Del latín ·tubus·. 1. m. Pieza hueca, de forma por lo común cilíndrica
y generalmente abierta por ambos extremos.
·Tubular:
rae; Del latín ·tubülus· 'tubo pequeño' y -ar. 1. adj. Peteneciente o relativo al tubo.
2.adj. Que tiene forma de tubo. 3. adj. Que está formado por tubos. 4. m. Cubierta de
una rueda de bicicleta en forma de tubo, com cámara de aire en su interior.
·Turba:
rae; Del francés ·tourbe·, y este del franco *turba; confer alto alemán antiguo
·zurba·, inglés antiguo ·turf·, nórdico ·torf·. 1. f Carbón fósil formado de
residuos vegetales, de color pardo oscuro, aspecto terroso y poco peso. 2. f. Estiércol
mezclado con carbón mineral que se emplea como combustible en los hornos de ladrillos.
·Turbera:
rae; 1. f. Sitio donde yace la turba.